Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Stemcorp.com.br
Stem Cell Rev and Rep (2010) 6:560–566DOI 10.1007/s12015-010-9187-5
Human Multipotent Mesenchymal Stromal Cellsfrom Distinct Sources Show Different In Vivo Potentialto Differentiate into Muscle Cells When Injectedin Dystrophic Mice
N. M. Vieira & E. Zucconi & C. R. Bueno Jr. & M. Secco &M. F. Suzuki & P. Bartolini & M. Vainzof & M. Zatz
# Springer Science+Business Media, LLC 2010
Abstract Limb-girdle muscular dystrophies are a hetero-
differentiate in muscle cells in vivo or if this capability is
geneous group of disorders characterized by progressive
influenced by the niche from where they were obtained. In
degeneration of skeletal muscle caused by the absence or
order to address this question we injected human derived
deficiency of muscle proteins. The murine model of
umbilical cord tissue MSCs (hUCT MSCs) into the caudal
Limb-Girdle Muscular Dystrophy 2B, the SJL mice,
vein of SJL mice with the same protocol previously used
carries a deletion in the dysferlin gene. Functionally, this
for hASCs; we evaluated the ability of these cells to engraft
mouse model shows discrete muscle weakness, starting at
into recipient dystrophic muscle after systemic delivery, to
the age of 4–6 weeks. The possibility to restore the
express human muscle proteins in the dystrophic host and
expression of the defective protein and improve muscular
their effect in functional performance. These results are of
performance by cell therapy is a promising approach for
great interest for future therapeutic application.
the future treatment of progressive muscular dystrophies(PMD). We and others have recently shown that human
Keywords Human multipotent mesenchymal stromal
adipose multipotent mesenchymal stromal cells (hASCs)
cells . Xenotransplantation . Muscular dystrophy . Therapy
can differentiate into skeletal muscle when in contact withdystrophic muscle cells in vitro and in vivo. Umbilicalcord tissue and adipose tissue are known rich sources of
multipotent mesenchymal stromal cells (MSCs), widelyused for cell-based therapy studies. The main objective of
Multipotent mesenchymal stromal cells (MSCs) are potentially
the present study is to evaluate if MSCs from these two
useful for therapeutic approaches as well as models for
different sources have the same potential to reach and
developmental biology studies. MSC can be isolated fromdifferent tissues, such as adipose tissue, dental pulp, placenta,umbilical cord and fallopian tube []. However an
N. M. Vieira and E. Zucconi contributed equally for this work.
important question is whether MSCs from different sourcesare comparable in their differentiation potential in vivo or
N. M. Vieira : E. Zucconi : C. R. Bueno Jr. : M. Secco :M. Vainzof : M. Zatz (*)
whether this potential is influenced by the niche from where
Human Genome Research Center, Institute of Biosciences,
Progressive muscular dystrophies (PMD) are a clinically
Rua do Matão, n.106—Cidade Universitária,
and genetically heterogeneous group of disorders caused by
São Paulo, SP CEP: 05508-090, Brazile-mail: mayazatz@usp.br
the deficiency or abnormal muscle proteins, resulting inprogressive degeneration and loss of skeletal muscle function.
As effective treatments for these diseases are still unavailable,
Biotechnology Department, National Nuclear Energy
they have been widely investigated as possible candidates for
Among the different forms, the Limb Girdle Muscu-
lar Dystrophies (LGMDs) constitute a sub-group char-acterized by the involvement of the pelvic and shouldergirdle musculature. A 171-bp in-frame deletion in themurine dysferlin cDNA was identified in a mousemodel, the SJL mice, with a corresponding reduction indysferlin levels to 15% of normal []. The SJL micedeletion is in-frame, and therefore does not cause a totalabsence of the protein.
The continuous and gradual muscle degeneration in
PMDs leads to a depletion of satellite cells and,consequently, the capability to restore the skeletalmuscle is lost [, ]. Different sources of stem/progenitorcells that show extended proliferation in vitro and alsohave the ability to generate normal muscle fibers in vitroand in vivo have been described in several publications , –].
We recently showed that human adipose-derived stro-
mal cells (hASCs) can differentiate into skeletal musclewhen in contact with dystrophic muscle cells in vitro and in vivo In addition we also observed that the humanumbilical cord tissue (hUCT) is a much richer source of
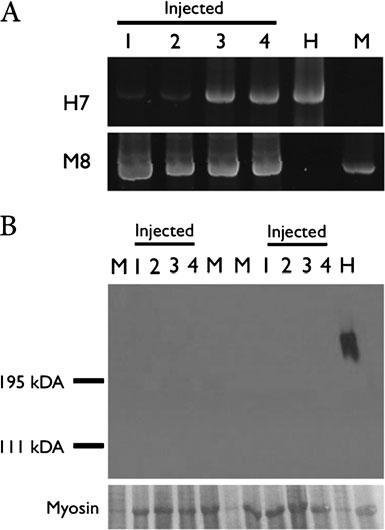
Fig. 1 a Polymerase chain reaction analysis for human chromosome 7α-satellite sequences (H7) and mouse chromosome 8 centromeric
MSC than umbilical cord blood and that they have
repeat sequence (M8) of SJL mice. Muscles of the injected SJL mice,
different expression profiles ]. However it is not known if
samples shown are the following: 1–Distal foreleg muscle; 2–
all MSCs show the same capacity in vivo. Do MSCs from
Proximal foreleg muscle; 3–Distal hindleg muscle; 4–Proximal hind-
adipose and umbilical cord tissue have the same potential to
leg muscle; H–Human DNA; M–Mouse DNA. b Western blot analysisfor human-dystrophin of the muscles of two injected animals. Samples
reach and differentiate into muscle cells in vivo? Or, this
shown are the following: 1–Distal foreleg muscle; 2–Proximal foreleg
capability is influenced by the niche from where they were
muscle; 3–Distal hindleg muscle; 4–Proximal hindleg muscle; H–
Human muscle protein; M–Mouse muscle protein. Myosin = myosin
In order to address this question we have injected hUCT
band detected in the Ponceau S pre-stained blot, for the evaluation ofloaded muscle proteins
MSCs intravenously into the SJL mice, aiming to comparetheir potential to differentiate into skeletal muscle with ourprevious data with hASCs []. Differently from hASCs,
Muscle Differentiation in the Host Muscle
hUCT MSCs reached the muscle but did not differentiateinto muscle cells. These results suggest that according to the
To explore the myogenic differentiation followed by the
source from which MSCs were obtained they may show a
engraftment of hUCT MSCs we analyzed the expression of
greater potential to differentiate into determined cell lineages.
dysferlin and human-dystrophin in the host muscle.
This may have important implications depending on the
The analysis of dysferlin is not sufficient to infer if the
injected muscles are expressing human or mouse proteins]. Therefore, we assessed the presence of human-dystrophin, using a specific anti-human-dystrophin anti-body [Through western blot (WB) analysis, no human
dystrophin was found in the muscles of the injected animals(Fig.
hUCT MSCs Capacity to Reach and Engraft at the HostMuscle
In order to assess the potential of hUCT MSCs to reach and
We performed three standardized motor ability tests
colonize the host muscle we injected undifferentiated,
and compared the performance of each SJL mouse
previously characterized, hUCT MSCs, into the caudal
(injected and uninjected) before (2-months of age) and
vein of SJL mice (n=7). PCR analysis detected human
after (9-months of age) the injections period, in blind test
DNA in the foreleg and hindleg muscles of all seven
(Table We observed that for the tests that required
trunk strength (inclined plane and wire hanging tests) the
Table 1 Results of 3 motor ability tests in injected (n =7) and uninjected mice (n=7) before and after 6 months of injection
At the inclined plane test the uninjected animals worsened their performance (p=0.008, t-Student test, n=7) while in the injected animals it did notdiffer (p=0.33, t-Student test, n=7)
For the wire hanging test the uninjected animals worsened their performance (p=0.0012, t-Student test, n=7) while the injected animals showedno significant difference (p=0.07, t-Student test, n=7)
At the ambulation test there was no difference in the performance of uninjected animals (p=0.11, t-Student test, n=7) and injected animals (p=0.16, t-Student test, n=7) after the injection period
uninjected animals showed a significantly worse perfor-
mice (30–90%) ]. Since the immunosupressive drug
mance while in the injected animals there were no
efficiently controlled the humoral and cellular immune
statistically significant changes (Table The deambula-
reactions, the authors concluded that the immune rejection
tion test did not show a significant difference before and
is not the cause of the low myoblast transplantation success
after the injections period in both groups.
We have previously shown that systemic delivery of
hASCs into the SJL mice, without immunosuppression,
resulted in human muscle proteins expression in the hostmuscles and functional amelioration [
The successful use of stem cells for clinical applications in
MSCs may be found in different tissues which are
therapy for PMD requires the finding of a rich and easily
obtainable source of cells, which must have the ability to
important question to be addressed is whether stem-cells
reach the entire body musculature, engraft and restore the
obtained from different sources have the same potential to
defective protein in the dystrophic muscle.
differentiate into different cell lineages or if there is already
Sampaolesi et al (2006) ] reported that systemic
a pre-commitment depending on the niche from which they
injections of normal dog mesoangioblasts to the muscle of
dystrophic dogs resulted in the restoration of dystrophin
Since umbilical cord is a rich source of MSCs, we
expression. However all transplanted dogs were maintained
investigated their ability to originate muscle proteins and
on steroids and received immunosuppressant drugs, which
ameliorate functional parameters using the same animal
makes difficult to evaluate functional results, since it is
model and methodology proved to be successful in our
known that immunosuppressive and anti-inflammatory
drugs can ameliorate the phenotype in muscular dystrophy
DNA analysis showed that the hUCT MSCs were able to
reach the host muscle through systemic delivery. However
Leriche-Guérin et al. (2002) ] investigated the effect
we did not find human muscle proteins in the same muscle
of myoblast transplantation into the SJL mice muscle with
samples where the human DNA was present.
immunosupression. The percentage of dysferlin positive
The functional ability in the previous and current study
labeled fibers obtained in their study was lower than the
was evaluated by standardized motor ability tests [
percentage of dystrophin-positive fibers usually observed
]. However, for the SJL model, the most affected muscles
following the transplantation of normal myoblasts in mdx
are the ones that are responsible for trunk strength [].
In opposition to our previous study with hASCs [] the
In short, here we compared, for the first time, the ability
injected animals with hUCT MSCs did not show clinical
of MSCs obtained from human umbilical cord tissue and
improvement, but, surprisingly, the performance of non-
adipose tissue to engraft into recipient dystrophic muscle
injected animals was significantly worse than in the “treated”
after systemic delivery; express human muscle proteins in
animals. The mice from the latter group maintained their
the dystrophic host and their effect in functional perfor-
performance at the end of the injection period, in particular
mance using the same animal model and protocol. Our
for the wire hanging test, which requires most trunk strength,
results showed that although umbilical cord MSCs appar-
suggesting an apparent stabilization of the dystrophic
ently do not have the same potential to differentiate in
process. That is, even without differentiating in muscle cells,
human muscle proteins in vivo as hASCs they were able to
the injected hUCT MSCs may have a positive effect when
reach the muscle and showed an apparent therapeutic
interacting with the host muscle. Indeed there are growing
benefit in injected animals as compared to the control
evidences in the literature describing the immunosuppressive
group, probably due to their immunomodulatory effect.
properties of MSCS [Inflammatory infiltration is
The present investigation suggests that although MSC from
observed in the dystrophic muscle but little is known about
different sources show apparently similar properties in vitro
the mechanisms involved in mesenchymal immunomodula-
they may be more or less efficient to differentiate into specific
tion. It is possible that secreted known cytokines factors
cell lineages in vivo according to the niche from where they
(TNF-α, IFN-γ and IL-12) could act, by protecting the
were obtained. Preclinical studies in different animal models,
dystrophic muscle. Several authors showed that mesenchy-
which are currently underway, will be essential to corroborate
mal stem cells suppress proliferation of activated lympho-
the present observations, which will have important implica-
cytes in vitro in a dose-dependent, non-HLA-restricted
tions aiming future cell therapy replacement.
manner [–]. Antibody-mediated depletion of CD4+and CD8+ T cells in mdx mice has been found to result ina reduction in muscle pathology []. MSCs are also being
tested in clinical trials aiming to ameliorate graft-versus-hostdisease after haemopoietic-stem-cell transplantation in
humans ]. Therefore, the immunomodulation effect ofMSCs in patients affected by progressive muscular dystro-
This study was approved by the human research ethics
phies could be a promising additional benefit to cell therapy.
committee (Comitê de ética em pesquisa—seres humanos—
Although MSCs from different sources show similar ability
CEP) and by the animal research ethics committee (Comissão
to differentiate into muscle cells in vitro [, , ] preclinical
de ética no uso de animais em experimentação—CEUA) of
studies are of utmost importance to verify it this also happens
Institute of Bioscience and University Hospital of University
in vivo. The apparent greater potential of adipose tissue than
of São Paulo. hUCT MSCs were collected from donated
umbilical cord derived MSCs to differentiate into muscle
umbilical cord units (UC), after all mothers signed the writhen
cells here observed could be explained by a recently
informed consent, in accordance with the ethical committee of
described population of mesenchymal progenitors, distinct
Institute of Bioscience and University Hospital of University
from satellite cells, in the skeletal muscle These
of São Paulo (CEP), permit number 040/2005. SJL mice were
progenitors have many similarities with hASCs and accord-
purchased from the Jackson Laboratory. Animal care and
ing to the authors they may have the same origin. These cells
experiments were performed in accordance with the animal
do not generate myofibers but enhance the rate of differen-
research ethics committee (CEUA) of the Biosciences
tiation of primary myogenic progenitors, and have adipo-
Institute, University of São Paulo, permit number 034/2005.
genic differentiation potential both in vitro and in vivo. Theinteraction between muscle cells and these mesenchymal
progenitors has a considerable impact on muscle homeostasissince adipogenesis is strongly inhibited by the presence of
UCs were filled with 0.1% collagenase (Sigma-Aldrich,
satellite cell-derived myofibres ]. It remains unclear
however which cell population participates in the regenera-
incubated at 37°C for 20 min. Each UC was washed with
tion process by fusing to the degenerated myotubes or
proliferation medium (DMEM low glucose, 10% fetal
forming new myofibers. The identification of this sub-
bovine serum), and the detached cells were harvested after
population will be extremely important for the establishment
gentle massage of the UC. Cells were centrifuged at 300 g
of clinical trial protocols. Interestingly, it has been recently
for 10 min, resuspended in proliferation medium, and
shown that there is an epigenetic memory in induced
seeded in 25-cm2 flasks at a density of 5 × 107 cells per ml.
pluripotent stem-cells according to the tissue of origin
After 24 h of incubation, non-adherent cells were removed
which might occur also with adult MSCS derived cells.
were fixed with 4% paraformaldehyde for 30 min, washed, andstained with a working solution of 0.16% oil red O for 20 min.
To analyze cell-surface expression of specific markers,adherent cells were incubated with the following anti-human
primary antibodies: CD29-PECy5, CD34-PerCP, CD31-phycoerythrin (PE), CD45-fluorescein isothiocyanate (FITC),
A pellet culture system was used for chondrogenesis.
CD90-R-PE, CD73-PE, CD13-PE, CD44-PE, CD117-PE,
Cells (2,5×105) were centrifuged in a 15-ml polypropylene
human leukocyte antigen (HLA)-ABC-FITC, HLA-DR-R-PE
tube at 500 g for 5 min, and the pellet was resuspended in
(Becton, Dickinson and Company, Franklin Lakes, NJ,
10 ml of basal medium consisting of DMEM-LG supple-
). A total of 10,000 labeled cells were analyzed
mented with 100 nM dexamethasone, 50 μM ascorbic acid-2
using a Guava EasyCyte flow cytometer running Guava
phosphate (Sigma-Aldrich), 1 mM sodium pyruvate
ExpressPlus software (Guava Technologies Hayward, CA,
(Invitrogen-Gibco), and 1% ITS-Premix (Becton Dickinson).
Without disturbing the pellet, cells were resuspended in 0.5 mlof chondrogenic differentiation medium consisting of basal
medium supplemented with 10 ng/ml transforming growthfactor-B1 (R&D Systems Inc., Minneapolis,
The evaluation of MSCs properties included immunopheno-
). On day 1, tubes were flipped gently to
typing by flow cytometric analysis, using a panel of surface
acquire a single floating cell sphere. Medium was changed
markers. hUCT MSCs were negative for CD31 (endothelial
every 3-4 days, and cells were fixed on day 21 with 4%
cell marker), CD34, CD45, CD117 (hematopoietic cell
paraformaldehyde. Cryosections (10 um thick) were stained
markers), and HLA-DR (human leukocyte differentiation
with toluidine blue to demonstrate extracellular matrix
antigen class II), whereas they were positive for CD29,
CD44 (adhesion markers), CD90, CD73, CD13 (mesenchy-mal markers), and HLA-ABC (human leukocyte differentia-
tion antigen class I) [] (data not shown).
The plasticity of hUCT MSCs was assessed by in vitro
To promote osteogenic differentiation, subconfluent cells
differentiation capacity, after three weeks of lineage
were treated with proliferation medium supplemented with
induction Myogenic, adipogenic, chondrogenic and
50 μM ascorbate-2 phosphate, 10 mM B-glycerophosphate
osteogenic differentiation was demonstrated by the expres-
(Sigma-Aldrich) and 0.1 μM dexamethasone, for 21 days.
sion of myogenic markers (myosin and desmin), lipid
Osteogenesis was demonstrated by accumulation of mineral-
vacuoles, mucopolysaccharide-rich extracellular matrix and
ized calcium phosphate assessed by von Kossa stain. Briefly,
calcium deposits, respectively. These results confirmed the
cells were stained with 1% silver nitrate (Sigma-Aldrich) for
mesenchymal nature of the isolated cells as well as their
45 min under ultraviolet light, followed by 3% sodium
multipotent potential (data not shown).
thiosulfate (Sigma-Aldrich) for 5 min, and then counterstainedwith van Gieson stain.
To evaluate MSCs properties, hUCT MSCs (third passage,at 80%–90% confluence) were subjected to adipogenic,
Fourteen two-months SJL mice were divided into two groups
chondrogenic, myogenic, and osteogenic differentiation in
of 7: transplanted animals (group A) and control group B
vitro, according to established protocols []. Normal human
(uninjected animals). Each animal from group A was injected
dermal fibroblasts were used as a negative control in the
in the tail vein with 1 ✕ 106 of hUCT MSC in 0.1 ml of
Hank’s Buffered Salt Solution (HBSS). The animals wereinjected for 6 months, weekly in the first month and then
monthly. All results were analyzed blindly. The code foreach of the mice groups was disclosed only after completion
Subconfluent cells were cultured in proliferation medium
of all the studies. Two months after the last cell transplan-
supplemented with 1 μM dexamethasone (Sigma-Aldrich),
tation, the animals were euthanatized using a CO2 chamber.
500 μM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), 60 μMindomethacin (Sigma-Aldrich), and 5 μg/ml insulin (Sigma-
Aldrich). Adipogenic differentiation was confirmed on day 21by intracellular accumulation of lipid-rich vacuoles stainable
The DNA was obtained using DNeasy Blood & Tissue Kit
with oil red O (Sigma-Aldrich). For the oil red O stain, cells
(Qiagen). The presence of human DNA in the host samples
were evaluated as described in Pelz et al (2005) ].
significance level of p=0.05 and the results were expressed
Centromeric region of human chromosome 7 and mice
by the percentage variation between their performance
chromosome 8 was amplified by PCR (35 cycles, annealing
before and after hUCT MSCs transfer period.
at 59°C). The PCR products were separated by electropho-resis on 2% agarose gels and stained with ethidium
Marcos Valadares, Tatiana Jazedje, Amanda Assoni, Mayra Pelatti,
bromide. Non-saturated digital images were obtained using
Juliana Gomes, Gabriela Polster, Camila Almeida, Agnes Nishimura,
an ImageQuant imaging system (GE HealthCare).
Natale Cavaçana, Miguel Mitne-Neto, Monize Lazar, Constancia Urbani,David Schlesinger, Daniela Bueno, Roberto Fanganiello, Antonia M P
Cerqueira, Marta Canovas, Paula Onofre and Dr. Maria Rita Passos-Bueno for helpful suggestions. We thank Dr. Glenn Morris from theCenter for Inherited Neuromuscular Disease (CIND), RJAH Orthopaedic
Muscle sample proteins were extracted through treatment with
Hospital, Oswestry, Shropshire, UK for providing anti-human dystrophin
a buffer containing 10 mM Tris-HCl (pH 8.0), 150 mM NaCl,
5 mM EDTA, 1% Triton X-100 and 60 mM octylglucoside. Samples were centrifuged at 13,000 × g for 10 min to removeinsoluble debris. Soluble proteins were resolved by 6%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE), and transferred to nitrocellulose membranes
1. Secco, M., Zucconi, E., Vieira, N. M., et al. (2008). Multipotent
(Hybond; Amersham). All membranes were stained with
stem cells from umbilical cord: cord is richer than blood! StemCells, 26, 146–50.
Ponceau (Sigma) to evaluate the amount of loaded
2. Zuk, P. A., Zhu, M., Mizuno, H., et al. (2001). Multilineage cells
proteins. Blots were blocked for 1 h in Tris-buffered
from human adipose tissue: implications for cell-based therapies.
saline Tween (TBST) containing 5% powdered skim milk
and reacted overnight with the following primary
3. Gang, E. J., Jeong, J. A., Hong, S. H., et al. (2004). Skeletal
myogenic differentiation of mesenchymal stem cells isolated from
antibody: anti human-dystrophin MANEX 12/16E2 G10
human umbilical cord blood. Stem Cells, 22, 617–24.
(1:100) kindly provided by Dr. Glenn E. Morris at
4. Gronthos, S., Brahim, J., Li, W., et al. (2002). Stem cell properties of
Center for Inherited Neuromuscular Diseases, Oswestry,
human dental pulp stem cells. Journal of Dental Research, 81, 531–5.
Shropshire, UK. Blots were incubated one hour with
5. Lee, O. K., Kuo, T. K., Chen, W. M., Lee, K. D., Hsieh, S. L., &
Chen, T. H. (2004). Isolation of multipotent mesenchymal stem
secondary antibodies. Immunoreactive bands were
cells from umbilical cord blood. Blood, 103, 1669–75.
detected with ECL chemiluminescence detection system
6. Jazedje, T., Perin, P. M., Czeresnia, C. E., et al. (2009). Human
fallopian tube: a new source of multipotent adult mesenchymalstem cells discarded in surgical procedures. Journal of Transla-tional Medicine, 7, 46.
7. Bittner, R. E., Anderson, L. V., Burkhardt, E., et al. (1999).
Dysferlin deletion in SJL mice (SJL-Dysf) defines a natural
In order to verify whether injected hUCT MSCs would
model for limb girdle muscular dystrophy 2B. Nature Genetics,
improve motor ability in SJL injected mice, we performed
8. Heslop, L., Morgan, J. E., & Partridge, T. A. (2000). Evidence for
motor ability tests before and after 6 months of SC injection
a myogenic stem cell that is exhausted in dystrophic muscle.
period. Mice were examined, weighed, and submitted to the
Journal of Cell Science, 113(Pt 12), 2299–308.
following tests: (a) the inclined plane test evaluated by
9. Laguens, R. (1963). Satellite cells of skeletal muscle fibers in
measuring the maximal angle of a wood board on which the
human progressive muscular dystrophy. Virchows Archiv fürPathologische Anatomie und Physiologie und für Klinische
animal was placed until it slipped; (b) the wire hanging test
to determine the ability of the mouse suspended on a
10. Gronthos, S., Mankani, M., Brahim, J., Robey, P. G., & Shi, S.
horizontal thread by its forelegs, to reach it with its hindlegs
(2000). Postnatal human dental pulp stem cells (DPSCs) in vitro
and the length of time they were able to stay hanging; (c)
and in vivo. Proceedings of the National Academy of Sciences ofthe United States of America, 97, 13625–30.
the ambulation test which was performed to determine the
11. Gussoni, E., Soneoka, Y., Strickland, C. D., et al. (1999).
mean length of a step measured in hindfoot ink prints while
Dystrophin expression in the mdx mouse restored by stem cell
mice freely run in a corridor (length, 50 cm; width, 8 cm;
transplantation. Nature, 401, 390–4.
12. Sampaolesi, M., Blot, S., D’Antona, G., et al. (2006). Meso-
angioblast stem cells ameliorate muscle function in dystrophicdogs. Nature, 444, 574–9.
13. Chan, J., Waddington, S. N., O’Donoghue, K., et al. (2007).
Widespread distribution and muscle differentiation of human fetal
Observations were quantified blindly. Numerical data are
mesenchymal stem cells after intrauterine transplantation indystrophic mdx mouse. Stem Cells, 25, 875–84.
the mean sd (standard deviation). The statistical analysis of
14. Kong, K. Y., Ren, J., Kraus, M., Finklestein, S. P., & Brown, R.
the equivalence between the injected and uninjected mice
H., Jr. (2004). Human umbilical cord blood cells differentiate into
was achieved by the one-tailed t-student test, at the
muscle in sjl muscular dystrophy mice. Stem Cells, 22, 981–93.
15. Vieira, N. M., Brandalise, V., Zucconi, E., et al. (2008). Human
ataxia reveal autophagic neurodegeneration in dorsal root ganglia.
multipotent adipose-derived stem cells restore dystrophin expres-
The Journal of Neuroscience, 24, 1987–95.
sion of Duchenne skeletal-muscle cells in vitro. Biology of the
27. Yonemori, F., Yamaguchi, T., Yamada, H., & Tamura, A. (1998).
Evaluation of a motor deficit after chronic focal cerebral ischemia
16. Vieira, N. M., Bueno, C. R., Jr., Brandalise, V., et al. (2008).
in rats. Journal of Cerebral Blood Flow and Metabolism, 18,
SJL dystrophic mice express a significant amount of human
muscle proteins following systemic delivery of human adipose-
28. Uccelli, A., Moretta, L., & Pistoia, V. (2008). Mesenchymal
derived stromal cells without immunosuppression. Stem Cells,
stem cells in health and disease. Nature Reviews. Immunology,
17. Secco, M., Zucconi, E., Vieira, N. M., et al. (2008). Mesenchymal
29. Klyushnenkova, E., Mosca, J. D., Zernetkina, V., et al. (2005). T
stem cells from umbilical cord: do not discard the cord!
cell responses to allogeneic human mesenchymal stem cells:
Neuromuscular Disorders, 18, 17–8.
immunogenicity, tolerance, and suppression. Journal of Biomed-
18. Secco, M., Moreira, Y. B., Zucconi, E., et al. (2009). Gene
expression profile of mesenchymal stem cells from paired
30. Le Blanc, K., Tammik, L., Sundberg, B., Haynesworth, S. E., &
umbilical cord units: cord is different from blood. Stem Cell
Ringden, O. (2003). Mesenchymal stem cells inhibit and stimulate
mixed lymphocyte cultures and mitogenic responses independent-
19. Thanh, L. T., Nguyen, T. M., Helliwell, T. R., & Morris, G. E.
ly of the major histocompatibility complex. Scandinavian Journal
(1995). Characterization of revertant muscle fibers in Duchenne
muscular dystrophy, using exon-specific monoclonal antibodies
31. Bartholomew, A., Sturgeon, C., Siatskas, M., et al. (2002).
against dystrophin. American Journal of Human Genetics, 56,
Mesenchymal stem cells suppress lymphocyte proliferation in
vitro and prolong skin graft survival in vivo. Experimental
20. Kennel, P. F., Fonteneau, P., Martin, E., et al. (1996). Electro-
myographical and motor performance studies in the pmn mouse
32. Spencer, M. J., Montecino-Rodriguez, E., Dorshkind, K., &
model of neurodegenerative disease. Neurobiology of Disease, 3,
Tidball, J. G. (2001). Helper (CD4(+)) and cytotoxic (CD8(+)) T
cells promote the pathology of dystrophin-deficient muscle.
21. Davies, K. E., & Grounds, M. D. (2006). Treating muscular
dystrophy with stem cells? Cell, 127, 1304–6.
33. Le Blanc, K., Frassoni, F., Ball, L., et al. (2008). Mesenchymal
22. Leriche-Guerin, K., Anderson, L. V., Wrogemann, K., Roy, B.,
stem cells for treatment of steroid-resistant, severe, acute graft-
Goulet, M., & Tremblay, J. P. (2002). Dysferlin expression after
versus-host disease: a phase II study. Lancet, 371, 1579–86.
normal myoblast transplantation in SCID and in SJL mice.
34. Jazedje, T., Secco, M., Vieira, N. M., et al. (2009). Stem cells
Neuromuscular Disorders, 12, 167–73.
from umbilical cord blood do have myogenic potential, with and
23. Partridge, T. A., Morgan, J. E., Coulton, G. R., Hoffman, E. P., &
without differentiation induction in vitro. Journal of Translational
Kunkel, L. M. (1989). Conversion of mdx myofibres from
dystrophin-negative to -positive by injection of normal myoblasts.
35. Uezumi, A., Fukada, S., Yamamoto, N., Takeda, S., & Tsuchida,
K. (2010). Mesenchymal progenitors distinct from satellite cells
24. Zucconi, E., Vieira, N. M., Bueno, D. F., et al. (2009).
contribute to ectopic fat cell formation in skeletal muscle. Nature
Mesenchymal stem cells derived from canine umbilical cord
vein—a novel source for cell therapy studies. Stem Cells and
36. Joe, A. W., Yi, L., Natarajan, A., et al. (2010). Muscle injury
activates resident fibro/adipogenic progenitors that facilitate myo-
25. Groshong, J. S., Spencer, M. J., Bhattacharyya, B. J., et al. (2007).
genesis. Nature Cell Biology, 12, 153–63.
Calpain activation impairs neuromuscular transmission in a mouse
37. Kim, K., Doi, A., Wen, B., et al. (2010). Epigenetic memory in
model of the slow-channel myasthenic syndrome. Journal of
induced pluripotent stem cells. Nature.
Clinical Investigation, 117, 2903–12.
38. Pelz, O., Wu, M., Nikolova, T., et al. (2005). Duplex polymerase
26. Simon, D., Seznec, H., Gansmuller, A., et al. (2004). Friedreich
chain reaction quantification of human cells in a murine
ataxia mouse models with progressive cerebellar and sensory
background. Stem Cells, 23, 828–33.
Lenisfera Dr Saronni: i benefici della naturaL’Echinacea (echinacea angustifolia) è una pianta erbacea perenneoriginaria del Nord America che usata esternamente ha proprietà cicatrizzante, riepitelizzante, antinfiammatoria, antisettica, antibatterica, decongestionante. L’Echinacea, ad uso cosmetico è indicata per pellisecche, aride, screpolate, e utile come decongestionante, rassodan
ASUNTOHAKEMUS HAKIJAN HENKILÖTIEDOT Sukunimi ja entiset nimet (painokirjaimin) Haluatko ilmoituksen myös sähköpostina AVIO/AVOPUOLISON HENKILÖTIEDOT (täytetään vain jos puoliso muuttaa haettavaan asuntoon) MUUT ASUMAAN TULEVAT HENKILÖT (tarvittaessa käytettävä liitettä) HAETTAVA HUONEISTO Muita toivomuksia (esim. vuokran suuruusluokka) ASUNNON TARVE (k
 Among the different forms, the Limb Girdle Muscu-
lar Dystrophies (LGMDs) constitute a sub-group char-acterized by the involvement of the pelvic and shouldergirdle musculature. A 171-bp in-frame deletion in themurine dysferlin cDNA was identified in a mousemodel, the SJL mice, with a corresponding reduction indysferlin levels to 15% of normal []. The SJL micedeletion is in-frame, and therefore does not cause a totalabsence of the protein.
Among the different forms, the Limb Girdle Muscu-
lar Dystrophies (LGMDs) constitute a sub-group char-acterized by the involvement of the pelvic and shouldergirdle musculature. A 171-bp in-frame deletion in themurine dysferlin cDNA was identified in a mousemodel, the SJL mice, with a corresponding reduction indysferlin levels to 15% of normal []. The SJL micedeletion is in-frame, and therefore does not cause a totalabsence of the protein.