Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
120295.qxd
S.K. Naess · J.M. Bradeen · S.M. Wielgus G.T. Haberlach · J.M. McGrath · J.P. Helgeson
Resistance to late blight in Solanum bulbocastanum is mappedto chromosome 8
Received: 26 November 1999 / Accepted: 22 December 1999
Abstract Somatic hybrids between potato and Solanum
There are several sources of late blight resistance
bulbocastanum, a wild diploid (2n=2x=24) Mexican spe-
available to the potato breeder, particularly among Mexi-
cies, are highly resistant to late blight, caused by Phyt-
can Solanum species which co-evolved with Phytophth-ophthora infestans. Both randomly amplified polymor-
ora infestans (van Soest et al. 1984). Solanum demissum,
phic DNA (RAPD) and restriction fragment length poly-
a Mexican hexaploid species, has been used extensively
morphism (RFLP) markers that are closely linked to the
in potato breeding programs. Much of the resistance to
resistance have been noted by analysis of three different
late blight in this species is due to a series of race-specific
backcross-2 populations derived from two different so-
resistance (R) genes. Unfortunately, the pathogen is able
matic hybrids. With reference to previously published
to overcome this resistance when presented with the few
potato and tomato maps, resistance appears to be on the
R genes introgressed into the genome of present-day
long arm of chromosome 8 and is flanked by RFLP
markers CP53 and CT64. In a population of BC plants
A potentially more durable race-non-specific re-
line J10lK6 sistance is present in many of the diploid Mexican spe-
[(S. tuberosum PI 203900+S. bulbocastanum PI 243510)
cies, notably S. bulbocastanum (Neiderhauser and Mills
×Katahdin)]×Atlantic, late blight resistance cosegregated 1953). These species are largely sexually incompatiblewith RFLP marker CT88 and RAPD marker OPG02–625.
with potato due to differences in endosperm balancenumbers (EBN, Johnston et al. 1980). Ploidy manipula-
Key words Solanum bulbocastanum · Late blight ·
tions and a series of bridge crosses have been success-
Phytophthora infestans · Somatic hybrid
fully used to effect hybridization between potato and S. bulbocastanum (Hermsen and Ramanna 1969; 1973;
Ramanna and Hermsen 1971; Hermsen and De Boer1971). However, reconstituting the recurrent potato par-
Late blight resistance is once again demanding the atten-
ent from the resulting quadruple species hybrids has
tion of potato breeders worldwide following recent
been difficult and late blight-resistant cultivars have not
migrations of aggressive metalaxyl-resistant isolates of
Phytophthora infestans into potato production areas.
More recently, somatic hybridization has been used to
Crop failures, reduced yields, storage losses, and in-
overcome sexual incompatibility between potato and
creased costs of crop protection, both to the grower and
S. bulbocastanum (Austin et al. 1993; Thieme et al.
to the environment, have resulted from these migrations
1997). Late blight resistance has been recovered in hexa-
(Fry and Goodwin 1997). Because no widely grown U.S.
ploid somatic hybrids and has been passed on undimin-
potato cultivar has adequate resistance to late blight, the
ished to progeny of the first and second backcrosses to
development of high-quality, late blight-resistant potato
potato (Helgeson et al. 1998; Douches et al. 1997). In
this paper we describe the mapping of late blight resis-tance from S. bulbocastanum to chromosome 8.
S.K. Naess · J.M. Bradeen · S.M. Wielgus · G.T. Haberlach
USDA/ARS Plant Disease Resistance Research Unit,
Department of Plant Pathology, University of Wisconsin,Madison, WI 53706, USA
Plants examined in these experiments were BC and BC proge-
nies descendant from somatic fusion products between Solanumbulbocastanum PI 243510 (2n=2x=24) and S. tuberosum, PI
23900 (2n=4x=48) (Helgeson et al. 1998). For convenience, the
gel (Diversified Biotech, Boston, Mass.) 0.6% agarose gel, stained
clones of S. bulbocastanum and S. tuberosum used in this study
with ethidium bromide and visualized on a UV transilluminator.
are designated PT29 and R4, respectively. The BC progenies
RAPD markers were grouped into synteny groups and the synteny
were obtained from crosses between three different somatic hy-
groups assigned to chromosomes with chromosome-specific RFLP
brids (J101, J103, and J138) and the S. tuberosum cultivars Katah-
probes as described in McGrath et al. 1996.
din (KAT), or Atlantic (ATL). Subsequently, BC progenies were
obtained from crosses between selected late blight-resistant BC1
individuals J101K6 and J101K27 and the potato cultivars ATL and
Norland (NOR), respectively. A third progeny group (J103K7
×A89804-7) was provided by Dr. Joe Pavek, Aberdeen, Idaho. DNA samples (10 µg) were digested with EcoRl, Hindlll, EcoRV,
These three BC populations are designated 1K6, 1K27 and LB1,
or Dral at a DNA/enzyme ratio of 7 units per microgram. The
fragments were size-separated on 0.8% agarose gels and blottedonto Hybond-N+ nylon membranes (Amersham Pharmacia Bio-tech, Piscataway, N.J.). Non-radioactive hybridization and detec-
tion methods were used following protocols provided by the man-ufacturer using either the ECL, Gene Images or AlkPhos systems
Plants were screened for late blight resistance in greenhouse facili-
(Amersham Pharmacia Biotech), depending on the probe used.
ties at the University of Wisconsin Biotron. Five replicates of
Probes were chromosome-specific tomato genomic and cDNA
clonally propagated, 20- to 30-cm plants were arranged randomly
probes (Tanksley et al. 1992; Bonierbale et al. 1988) as well as
on carts for whole plant assays. Plants were sprayed to run-off
potato genomic probes (Gebhardt et al. 1989, 1991). Enzyme/
with a fine mist of Phytophthora infestans sporangial suspension
probe combinations which gave useful polymorphisms between
prepared from US-8, type A2, Cornell standard ME 93-A2
the potato and S. bulbocastanum genomes are as follows:
(WEF#US930287) cultures maintained on rye A medium. Thesuspension contained approximately 30000 sporangia/ml and was
chromosome 1: TG301/HindIII, TG17/EcoRI, TG27/EcoRI;
pre-chilled 4 h at 10°C before use. Relative humidity in the green-
chromosome 2: TG31/EcoRV, CT75/EcoRV, TG34/HindIII, GP504/
house was maintained at or above 90%. The temperature was
maintained at 23°C during daylight hours (15 h) and dropped to
chromosome 3: TG56/EcoRI, TG130/HindIII, TG134/EcoRI,
15°C at night. Foliage blight scores were recorded at 4–5, 7,
10–11, and 14–15 days. A blight scale, with 0 indicating a dead
chromosome 4: TG123/EcoRI, TG65/EcoRI, TG450/EcoRI;
plant and 9 no visible infection, was used to visually rate disease
chromosome 5: CD64/HindIII, TG23/HindIII, TG185/HindIII;
severity. The ratings and the ranges of percentage infections asso-
chromosome 6: TG231/EcoRI, CT83/HindIII, TG25/EcoRI,
ciated with the rating value are as follows: 9, no visible infection;
8, <10%; 7, 11–25%; 6, 26–40%; 5, 41–60%; 4, 61–70%; 3, 71–
chromosome 7: TG128/EcoRI, TG61/EcoRI, TG272/EcoRV;
80%; 2, 81–90% 1, >90%; 0, 100% (dead).
chromosome 8: TG176/EcoRI, GP245(GP170)/HindIII, TG45/
In preliminary field trials, late blight resistance segregated 1:1
DraI, TG41/EcoRV, GP171/DraI, CT245/HindIII,
in BC progeny of S. bulbocastanum+S. tuberosum somatic hy-
CP53/EcoRV, PPO/HindIII, TG495/EcoRV, CT88/
brids, indicating that late blight resistance in these plants may be
EcoRI, TG478/HindIII, CT64/HindIII, TG261/
due to a single dominant gene or a tightly linked cluster of genes. EcoRI, GP301/EcoRI, CT148/HindIII, CT265/
The late blight data were therefore recoded from quantitative to
EcoRI, CT252/EcoRI, CT68/EcoRI;
qualitative for mapping purposes (Table 1). BC clones with aver-
chromosome 9: TG18/EcoRV, TG35/HindIII, TG404/EcoRV,
age resistance scores of 8.0 or above (10% or less leaf infection)
were scored as resistant and those with a resistance score of 6.9 or
chromosome 10: TG122/HindIII, TG52/EcoRV, CT20/HindIII,
below (>25% infection) as susceptible. Clones with an average re-
sistance score of 7.0–7.8 were omitted from the initial analysis
chromosome 11: GP125/EcoRI, CT168/EcoRV, CT55/EcoRV,
(5/50, 3/54 and 2/69 individuals in the 1K27, 1K6 and LB1 popu-
lations, respectively. The recoded resistance data were included in
chromosome 12: TG68/HindIII, CT211/EcoRV, TG28/EcoRV,
the MAPMAKER random amplified polymorphic DNA (RAPD)
In addition, two RAPD fragments, G02-575 and P09-550, of S. bulbocastanum origin were cloned for use as RFLP probes (see
For RAPD analysis, DNA was extracted from single leaves of invitro-grown material by the miniprep method described in
Sequence characterized amplified region (SCAR) and cleaved
McGrath et al. (1994). A microprep DNA extraction protocol was
amplified polymorphic sequence (CAPS) markers
used when larger quantities of DNA were required for restrictionfragment length polymorphism (RFLP) analysis (Fulton et al.
The Original TA Cloning Kit (INVITROGEN, Carlsbad, Calif.)
was used to clone RAPD fragments of interest. RAPD fragmentswere first reamplified using a band-stab technique (Bjourson andCooper 1992), and 1-µl aliquotes of the reamplification reactions
were ligated into the pCR2.1 vector following instructions provid-ed by the manufacturer.
Amplification reactions were carried out in 25-µl reaction mix-
An ABI automatic sequencer (Perkin Elmer) was used to se-
tures containing 10 mM TRIS-HCl, pH 8.3, 50 mM KCl, 2 mM
quence plasmid inserts using fluorescent dye terminators. New
MgCl , 0.01% gelatin, 100 µM of each dNTP, 200 nM primer (OP-
primers were designed from sequences internal to the original
ERON, Alameda, Calif.), approximately 15 ng template DNA and
Operon primer annealing site. Primer sequences designed from the
1 Unit Amplitaq DNA Polymerase (Perkin Elmer, Foster City,
P09–550 fragment were AGG TGG TGG TGG GGT GGG ATA
Calif.) using a Perkin-Elmer model 480 thermocycler. The cycling
GTG and GTG GTC CGC ACC CAT ATT TCA CCA.
program consisted of an initial 2 min denaturation step at 94°C,
For SCAR markers based on RFLP probes, primers were first
followed by 3 cycles of 94°C (1 min), 35°C (1 min), and 72°C designed based on the published probe sequences (Solgenes dat-(2 min) then by 32 cycles of 94°C (30 s), 35°C (30 s), 72°C (1
abase:http://probe.nalusda.gov:8300/cgi-bin/browse/solgenes). These
min), and a final 5-min extension step at 72°C (McGrath et al.
primers, 905/906 (GTT GGG CAG AAG AGC TAG/ TTG CCT
1996). Amplification products were size-separated on a 1% Syner-
TAG TCC CCA GAG) and CT64 forward/reverse (GAG GAG
AGA TTC TTG GAC/ TGA GGT TGA TAG TGG GTG), wereused to generate fragments from both fusion parents using lowstringency polymerase chain reaction (PCR) conditions (see be-low). The resulting monomorphic fragments were cloned as de-scribed. Sequence differences between the CT88 plasmid insertsderived from the two fusion parents (PT29 and R4) were used todesign new primers specific to the resistant parent, PT29 (CT88forward and reverse: TGA GGC TCT TGG CTT TCG/AAT CAATTG AGC ATC TTG AGA).
For amplification using locus-specific primers 905/906 and
CT88 forward/reverse the MgCl and dNTP concentrations were
increased to 4 mM and 300 µM, respectively. The annealing tem-perature used with primers 905/906 and CT64 forward/reversewas 55°C and for CT88 forward/reverse it was 60°C. The cyclingprogram consisted of an initial 2-min. denaturation step at 94°Cfollowed by 30 cycles of 94°C for 1 min., an annealing step (55°Cor 60°C) for 30 s., and an elongation step at 72°C for 1.5 min. These cycles were followed by a final 5-min extension cycle at72°C.
For amplification of SCAR marker P09-478 a step-down PCR
Fig. 1 Field resistance to late blight derived from Solanum bulbo-
cycling program was used. The annealing temperature in the
castanum under intense disease pressure in Toluca, Mexico. BC3
above cycling program was varied. In the initial 3 cycles it was
individual A9509-34 (J101K6A22 X A84118-3) is contrasted with
72°C, in the following 3 cycles, 70°C and in the final 25 cycles,
For CAPS markers, fragments generated following amplifica-
tion with CT88 primers 905/906 were cleaved in the same reaction
were included in the MAPMAKER RAPD mapping data
tube with 1 U TaqI for 1 h at 65°C. The products were size-
sets. Segregation ratios for resistance did not deviate sig-
separated and visualized as previously described.
nificantly from 1:1 in the 1K27 or 1K6 populations; re-spectively 58% and 59% of the individuals in these pop-
ulations were resistant. Both of these populations werederived from somatic hybrid J101. In the LB1 popula-
Markers were mapped using the haploid function of MAPMAKER
tion, derived from somatic hybrid J103, only 37% of the
(Lander et al. 1987). Only markers specific to S. bulbocastanum
individuals were resistant, a significant deviation from a
were scored and included in the mapping data set. In using thehaploid function for mapping in the BC we are assuming that
1:1 segregation ratio (0.01<P 1:1<0.05).
there is a single copy only of each S. bulbocastanum chromosome
Late blight resistance maps to chromosome 8 and is
in the BC parent due to preferential pairing of the S. bulbocast-
linked to RAPD marker G02-625 in all three BC popu-
anum chromosomes in the somatic hybrid. RAPD markers were
lations (Fig. 2). In the 1K6 population, resistance coseg-
grouped at an LOD of 3. Easily scored but unassigned markers
regates with G02-625 ( Fig. 3, Table 1), and in the 1K27
were recoded (presence of marker recorded as absence and viceversa) to test whether any of these markers were segregating in re-
population a single individual (1/50) is recombinant be-
pulsion to other synteny groups. Only markers that give readily
tween resistance and G02-625. There is 9% recombina-
distinguished polymorphisms with potato were included in the
tion between resistance and G02-625 in the LB1 popula-
tion; 5/69 susceptible seedlings have the marker and1/69 resistant seedling lacks the marker.
In all three populations, resistance maps to the end of
the RAPD marker synteny group. However, in the LB1population markers G02-575 and P09-550 segregate in
In preliminary field trials, late blight resistance segregat-
repulsion to the rest of the RAPD synteny group. These
ed 1:1 in BC progeny of S. bulbocastanum+S. tubero-
markers, when recoded, flank resistance. The increased
sum somatic hybrids, indicating that late blight resis-
recombination rate between G02-625 and resistance in
tance in these plants may be due to a single dominant
the LB1 population may be due to the presence of por-
gene or a tightly linked cluster of genes. Resistance
tions of both PT29 chromosome 8 homologues in the
could be passed on, apparently undiminished, to BC
(Helgeson et al. 1998) and BC (Fig. 1) populations. In-
spection of the preliminary RAPD data indicated thatresistance was associated with one synteny group, later
RAPD markers G02-575 and P09-550 are heterozygousin PT29 and segregate in the combined BC populations.
Assignment of resistance to chromosome 8
This is also true for a number of other chromosome 8
RAPD markers which segregate in the BC but are pres-
ent in all 3 resistant individuals chosen as parents for the
Late blight resistance was assessed in three different BC
BC populations. Late blight resistance also segregates
populations. Representative results for one of these pop-
in the BC and it is therefore possible to map resistance
ulations are given in Table 1. The recoded resistance data
in these populations in relation to other segregating
Table 1 Association of RAPD marker G02-625 with late blight
0, dead. Late blight scores were recoded to R (<10% defoliation)
resistance in the 1K6× Atlantic BC population. Resistance scores:
0, (11–25% defoliation) or S (>25% defoliation) for qualitative
9, no visible infection; 8, <10%; 7, 11–25%; 6, 26–40%; 5,
mapping purposes. RAPD marker G02-625 is scored as present
41–60%; 4, 61–70%; 3, 71–80%; 2, 81–90%; 1>90% defoliation;
Fig. 2 Association of late
blight resistance with RAPDmarker G02-625 on chromo-some 8 in three different BC
are linked with a dotted lineRESISTANCE RESISTANCE RESISTANCE
markers previously mapped in the three BC popula-
and 13% in the BC 1K27, 1K6, and LB1 populations.
Thus, there appears to be some repression of recombina-
For initial considerations, we have assumed that the 3
tion in the BC populations relative to the BC popula-
somatic hybrids are identical with respect to chromo-
some 8 and have combined data from all BC progeny.
In this combined group of 103 individuals, recombina-tion between resistance and G02-625 is 11.5%. P09-550
Expansion of the chromosome 8 molecular map
is linked in repulsion to resistance and, when recoded,
maps to the other side of resistance as it does in theLB1s (Fig. 4). Recombination between K19-675 and
Molecular marker coverage of S. bulbocastanum chro-
G02-625, 2 RAPD markers that are common to all popu-
mosome 8 was expanded through RFLP analysis of 64
lations examined, is 28% in the BC but only 26%, 17%,
BC individuals in the 1K6 population. For this, chromo-
P2 SH T1 BC1 T2 SEGREGATING BC2s Recombination RESISTANCE CT88 CT64 TG261 Fig. 3 Cosegregation of RAPD marker G02-625 with late blight re- sistance in the 1K6 BC population. R and S Respectively late blight
resistant and susceptible individuals, P1 Solanum bulbocastanumfusion parent PI 243510 clone PT29, P2 S. tuberosum fusion parent
Fig. 5 Combined RAPD, RFLP marker coverage of chromosome
PI 23900 clone R4, SH somatic hybrid J101, T1 S. tuberosum back-
8 based on 64 BC individuals from the cross between J101K6 and
cross parent Katahdin, BC1 BC parent J101K6, T2 S. tuberosum
Atlantic. RFLP markers and resistance are in bold
late blight resistance in the 1K6 population, as has been
Recombination
previously noted for RAPD marker G02-625. RFLPmarker P09 maps distal to resistance, as anticipated fromthe position of the recoded RAPD marker in the BC and
LB1 population. Recombination between resistance and
RFLP marker P09 in this population is 10%.
Most of the RFLP probes used in this study are toma-
to cDNA clones and have not been included on previous-ly published potato maps. Therefore, we turned to the to-
mato map (Tanksley et al. 1992) in addition to potatomaps for useful comparisons. Colinearity between toma-to chromosome 8 and S. bulbocastanum chromosome 8appears to be conserved with only a few differences.
TG176 which maps distal to TG41 in both the tomato
and potato chromosome 8 maps, maps to chromosome 1
RESISTANCE
in the 1K6 population as it does in another BC popula-
tion derived from a different S. bulbocastanum accession(Brown et al. 1996). CT245 maps to two different loca-
Fig. 4 Association of late blight resistance and RAPD marker
tions on the tomato chromosome 8; 1 marker cosegre-
G02-625 on chromosome 8 in the combined BC progeny. Marker
gates with TG176 and the other maps between TG41 and
name given by Operon primer followed by estimated marker size
PPO. In the 1K6 population, CT245 cosegregates with
in base pairs. Presumed allelic markers are linked with a dotted line
RFLP marker GP301 and is flanked by markers P09-2and CT148. CD40, which maps between RFLP markers
some 8-specific tomato cDNA and genomic probes,
TG176 and TG41 in tomato, cosegregates with CT252
potato genomic probes, and probes derived from RAPD
and CT68 at the other end of the chromosome in the 1K6
fragments were used. In this population late blight resis-
population. Finally, CT124, a probe specific for chromo-
tance cosegregates with RFLP marker CT88 and is
some 10 in tomato, hybridizes to several restriction frag-
flanked by RFLP markers CT64 and CP53 (Fig. 5).
ments in the 1K6 population, one of which maps to chro-
RAPD markers G02-575 and P09-550 from the LB1
mosome 10 and the other to chromosome 8. None of the
population were cloned and converted to RFLP probes
discrepancies between marker synteny or order involved
which hybridize to informative fragments in the 1K6
more than 1 consecutive marker. As all of these probes
population. RFLP marker G02-575 cosegregates with
hybridize to several restriction fragments in the 1K6
population it is possible that the fragments scored are
SCAR MARKER P09
non-orthologous loci and do not represent actual differ-
P1 P2 SH T1 BC1 T2 SEGREGATING BC2'S
ences in colinearity between tomato and S. bulbocast-
Recombination rates between markers common to the
tomato, potato, and 1K6 populations are lower in the1K6 population than in tomato but are comparable tothose in the potato map (Tanksley et al. 1992). The ge-netic distances between markers PPO and TG261, which
flank late blight resistance in the 1K6 population, are12.6 cM, 5.8 cM, and 2.6 cM, respectively, in the toma-
CAPS MARKER CT88
to, 1K6, and potato maps. In the 1K6 population the dis-tance between RFLP markers CT88 and CT64 is inflated
P1 P2 SH T1 BC1 T2 SEGREGATING BC2'S
relative to the tomato map (3.9 cM vs 1.4 cM), perhapsdue to the small size of the population examined. In anexpanded BC population (J101K6×NOR) of 273 indi-
viduals with the same maternal parent, recombintion be-tween CT88 and CT64 is only 0.8% (data not shown).
With the addition of RFLP markers to the RAPD map
it becomes clear that the RAPD markers are not random-ly distributed along the chromosome. Of the 14 RAPDmarkers mapped to chromosome 8, 11 map to an area
flanked by RFLP markers GP245 and TG41, an area of
Fig. 6 PCR-based markers generated from RAPD marker P09 and
only 13 cM in our map. Only 3 RAPD or RAPD-derived
RFLP marker CT88. In both panels P1 is the resistant (R) fusion
markers map to the region of chromosome 8 stretching
parent S. bulbocastanum PI243510 clone PT29, and P2 is the sus-
from RFLP marker TG41 to CT68, an area of 33 cM in
ceptible (S) fusion parent S. tuberosum PI203900 clone R4, SH so-
matic hybrid, T1 and T2 are, respectively, the S. tuberosum back-cross parents Katahdin and Atlantic, BC resistant backcross par-
ent J101K6. A SCAR marker P09, B CAPS marker CT88. Arrows mark the restriction products generated following digestion of the
PCR-based markers were designed from a RAPD markerlinked in repulsion to late blight resistance and from 2RFLP markers linked to resistance. Primers complemen-
reaction, in the PCR reaction mix, and the tubes were in-
tary to the sequence of RAPD marker P09-550 amplified
cubated at 65°C for 15 min prior to amplification. Only
a single fragment 478 bp in length which was specific to
the fragment specific to S. bulbocastanum is amplified
S. bulbocastanum (Fig. 6). These primers generate infor-
following digestion with TaqI.
mative markers in the 1K6 population, which lacksRAPD marker P09-550. They can be used to detect re-combination between RAPD marker G02-625 and SCAR
marker P09 which flank resistance in other populations.
Primers designed from the end sequences of tomato
The development of late blight-resistant potato varieties
probes CT64 and CT88 amplified monomorphic frag-
has re-emerged as a major objective of potato breeders
ments from S. bulbocastanum and the susceptible potato
around the world. Obstacles to sexual hybridization
fusion parent. The CT88 S. bulbocastanum fragment in-
between potato and the late blight resistant wild species
cludes a TaqI restriction site absent in the potato frag-
Solanum bulbocastanum have been overcome through
ment. Primers designed around nucleotide differences in
somatic hybridization, and late blight resistance from
the two sequences were not specific enough for the gen-
this species has been recovered in somatic hybrids, BC ,
eration of informative SCAR markers. We were much
BC , and BC progeny. This resistance appears to be race
more successful with CAPS for this marker, using the
non-specific; it provides sufficient resistance not only to
primers designed from the end sequences of the tomato
the US8 genotype of the pathogen but also to many races
probe, which flank the polymorphic TaqI site, to amplify
of Phytophthora infestans present in Toluca, Mexico
monomorphic fragments from all samples and cleaving
(Fig. 1) where numerous genotypes of the pathogen have
the resulting amplification product with TaqI (Fig. 6).
been isolated from resistant plants (Helgeson et al. 1998;
The CT64 fragments also contained a polymorphic
Grunwald personal communication). Nevertheless, much
TaqI restriction site. However, in this case the S. bulbo-
work remains to be done before introgression of late
castanum fragment lacks a site present in the S. tubero-
blight resistance from S. bulbocastanum into the potato
sum fusion and backcross parents. To take advantage of
genome is complete, and the goal of late blight resistant
this polymophism we included TaqI, at 1 U per 25 µl of
potato cultivars suitable to industry is realized.
In this study, genome coverage was sufficient to find
ever, PPO from S. bulbocastanum cannot be responsible
RAPD markers tightly linked to late blight resistance. By
for late blight resistance. We have found resistant indi-
mapping resistance in several different populations and
viduals lacking S. bulbocastanum-derived PPO genes
recoding unassigned markers to detect possible repulsion
and susceptible individuals with these genes. Leister et
phase linkages we were able to find 2 RAPD markers,
al. (1996) mapped numerous PCR-derived resistance
G02-625 and P09-550, which flank the resistance locus
gene analogs obtained from genomic potato DNA to the
12 potato chromosomes, including chromosome 8. Sola-
Late blight resistance mapped to the RAPD synteny
num tuberosum fragment St3.4e maps to the same 25-cM
group assigned to chromosome 8. Further expansion of
region, flanked by markers GP40 and GP36a, as the
the map with RFLP markers places resistance between
S. bulbocastanum-derived late blight resistance studied
RFLP markers CT64 and CP53. This area of chromo-
here. Although the sequence of many of the PCR-
some 8 has not previously been associated with late
derived probes obtained were related to sequences of
blight resistance in potato or tomato. Many mapping
known plant resistance genes, St3.4 did not show homo-
studies on late blight resistance in potato have been con-
logies to known genes (Leister et al. 1996). The St3.4
ducted on S. demissum-derived materials, and these have
fragment is multicopy in the potato genome and has as
indicated that a number of chromosomes are involved.
yet not been mapped in our populations. Whether or not
Several of the R genes from this species have been
St 3.4 is related to late blight resistance in S. bulbocast-
mapped, including R1 on chromosome 5 (Leonards-
Schippers et al. 1992), R2 on chromosome 4 (Li et
Suppression of recombination in introgression seg-
al. 1998) and R3, -6 and -7 on chromosome 11 (El
ments can lead to linkage drag persisting through many
backcross generations (Young and Tanksley 1989).
Quantitative trait loci (QTLs) associated with late
There is evidence of suppression of recombination in the
blight resistance have also been mapped in potato. These
BC populations examined as well as in a larger BC
may be more relevant to late blight resistance from
population ( preliminary data not shown). Marker-assisted
S. bulbocastanum, which is not governed by R genes
selection of parental material will therefore be crucial to
(Helgeson et al. 1998). Leonards-Schippers et al. (1994)
the success of the backcross program. Several late
found QTLs associated with late blight resistance on 9 of
blight-resistant individuals from the BC populations are
12 chromosomes in a potato mapping population segre-
missing all markers from 6 or more S. bulbocastanum
gating for race-non-specific resistance. None were found
chromosomes, a reduction in the wild species genome
on chromosome 8. Meyer et al. (1998) have also mapped
similar to that obtained through asymmetric somatic hy-
QTLs associated with late blight resistance in a potato
bridizations (Oberwalder et al. 1998). These individuals,
population segregating for race non-specific resistance.
as well as those resistant individuals recombinant for
Late blight resistance in their population was previously
chromosome 8, should be used to advantage as parental
reported to be located opposite the waxy locus on chro-
mosome 8 but has now been mapped to the short arm of
The generation of larger progeny populations will be
chromosome 4 near microsatellite STM 3016 ( De Jong,
necessary to compensate for the suppression of recombi-
personal communication). This marker is located be-
nation in the introgressed segments. Locus-specific
tween RFLP markers GP180 and GP172 (Milbourne et
PCR-based markers are needed to increase the efficiency
al. 1998), the same area to which a major QTL for late
of screening these progeny for those rare late blight-
blight resistance, Pi1a, maps in the Leonard-Schippers et
resistant individuals recombinant in the introgressed seg-
al. (1994) population. The race-non-specific tomato late
ment. SCAR and CAPS markers linked to late blight re-
blight resistance gene, Ph-2, maps to a single location on
sistance in S. bulbocastanum have been developed for
this purpose from 3 markers, P09–550, CT64 and CT88.
Late blight resistance in S. bulbocastanum does not
Acknowledgments We thank Dr. Joe Pavek, ARS/USDA
appear to be due to a large number of QTLs on different
Aberdeen, Idaho for the LB1 population and cross of
chromosomes. Quantitative resistance data from the
J101K6A22 with his breeding line A84118-3 and Dr.
J101K6×ATL BC population (Table 1) have been ana-
Hector Lozoya for the late blight testing of clone A9509-
lyzed using MAPMAKER QTL (Unix version 1.9) and a
nonparametric procedure (Mann-Whitney U test). Theseanalyses identified only one major chromosomal regionassociated with late blight resistance, the same area on
chromosome 8 previously identified by MAPMAKER(data not shown). Sixty-two percent of the observed vari-
Austin S, Pohlman JD, Brown CR, Mojtahedi H, Santo GS,
ability in disease resistance is explained by this chromo-
Douches DS, Helgeson JP (1993) Interspecific somatic hybrid-
ization between Solanum tuberosum L. and S. bulbocastanum
Several plant defense-related genes have been located
DUN. as a means of transferring nematode resistance. Am
on chromosome 8, including the PPO gene which is in-
Bonierbale MW, Plaisted RL, Tanksley SD (1988) RFLP maps
volved in Colorado potato beetle resistance in S. bertha-
based on a common set of clones reveal modes of chromosom-
ultii (Bonierbale et al. 1994). In our populations, how-
al evolution in potato and tomato. Genetics 120:1095–1103
Bonierbale MW, Plaisted RL, Pineda O, Tanksley SD (1994) QTL
Leister D, Ballvora A, Salamini F, Gebhardt C (1996) A PCR-
analysis of trichome-mediated insect resistance in potato.
based approach for isolating pathogen resistance genes from
potato with potential for wide application in plants. Nat Genet
Bjourson AJ, Cooper JE (1992) Band-stab PCR: a simple tech-
nique for the purification of individual PCR products. Nucleic
Li X, van Eck HJ, Rouppe van der Voort JNAM, Huigen DJ, Stam
P, Jacobsen E (1998) Autotetraploids and genetic mapping us-
Brown CR, Yang CP, Mojtahedi H, Santo GS, Masuelli R (1996)
ing common AFLP markers: the R2 allele conferring resis-
RFLP analysis of resistance to Columbia root-knot nematode
tance to Phytophthora infestans mapped on potato chromo-
derived from Solanum bulbocastanum in a BC population.
McGrath JM, Wielgus SM, Uchytil TF, Kim-Lee H, Haberlach
Douches DS, Kirk WW, Jastrzebski K, Long C, Hammerschmidt
GT, Williams CE, Helgeson HP (1994) Recombination of
R (1997) Susceptibility of potato varieties and advanced
Solanum brevidens chromosomes in the second generation
breeding lines (Solanum tuberosum L.) to Phytophthora infest-
from a somatic hybrid with S. tuberosum. Theor Appl Genet
ans (Mont.) De Bary in greenhouse screenings. Am Potato J
McGrath JM, Wielgus SM, Helgeson JP (1996) Segregation and
El-Kharbotly A, Palomino-Sanchez C, Salamini F, Jacobsen E,
recombination of Solanum brevidens synteny groups in proge-
Gebhardt C (1996) R6 and R7 alleles of potato conferring
ny of somatic hybrids with S. tuberosum: intragenomic equals
race-specific resistance to Phytophthora infestans (Mont.) de
or exceeds intergenomic recombination. Genetics 142:1335–
Bary identified genetic loci clustering with the R3 locus on
chromosome XI. Theor Appl Genet 92:880–884
Meyer RC, Milbourne D, Hackett CA, Bradshaw JE, McNichol
Fry WE, Goodwin SB (1997) Re-emergence of potato and tomato
JW, Waugh R (1998) Linkage analysis in tetraploid potato and
late blight in the United States. Plant Dis 81:1349–1357
association of markers with quantitative resistance to late
Fulton TM, Chunwongse J, Tanksley SD (1995) Microprep proto-
blight (Phytophthora infestans). Mol Gen Genet 259:150–160
col for extraction of DNA from tomato and other herbaceous
Milbourne D, Meyer RC, Collins AJ, Ramsay LD, Gebhardt C,
Waugh R (1998) Isolation, characterisation and mapping of
Gebhardt C, Ritter E, Debener T, Schachtschabel U, Walkemeier
simple sequence repeat loci in potato. Mol Gen Genet 259:
B, Uhrig H, Salamini F (1989) RFLP analysis and linkage
mapping in Solanum tuberosum. Theor Appl Genet 78:65–75
Moreau P, Thoquet P, Olivier J, Laterrot H, Grimsley N (1998)
Gebhardt C, Ritter E, Barone A, Debener T, Walkemeier B,
Genetic mapping of Ph-2, a single locus controlling partial re-
Schachtschabel U, Kaufmann H, Thompson RD, Bonierbale
sistance to Phytophthora infestans in tomato. Mol Plant –
MW, Ganal MW, Tanksley SD, Salamini F (1991) RFLP maps
of potato and their alignment with the homoeologous tomato
Neiderhauser JS, Mills WR (1953) Resistance of Solanum species
to Phytophthora infestans in Mexico. Phytopathology 43:
Helgeson JP, Pohlman JD, Austin S, Haberlach GT, Wielgus SM,
Ronis D, Zambolim L, Tooley P, McGrath JM, James RV,
Oberwalder B, Schilde-Rentschler L, Ruoss B, Wittemann S,
Stevenson WR (1998) Somatic hybrids between Solanum
Ninnemann H (1998) Asymmetric protoplast fusions between
bulbocastanum and potato: a new source of resistance to late
wild species and breeding lines of potato-effect of recipients
and genome stability. Theor Appl Genet 97:1247–1254
Hermsen JGTh, De Boer AJ (1971) The effect of colchicine treat-
Ramanna MS, Hermsen JGTh (1971) Somatic chromosome elimi-
ment on Solanum acaule and S. bulbocastanum; a complete
nation and meiotic chromosome pairing in the triple hybrid 6x-
analysis of ploidy chimeras in S. bulbocastanum. Euphytica
(Solanum acaule×S. bulbocastanum)×2x-S. phureja. Euphytica
Hermsen JGTh, Ramanna MS (1969) Meiosis in different F -
Soest LJM van, Schober B, Tazelaar MF (1984) Resistance to
hybrids of Solanum acaule Bitt. X S. bulbocastanum Dun and
Phytophthora infestans in tuber-bearing species of Solanum
it’s bearing on genome relationship, fertility and breeding
and it’s geographical distribution. Potato Res 27:393–411
Tanksley SD, Ganal MW, Prince JP, de Vicente MC, Bonierbale
Hermsen, JGTh, Ramanna MS (1973) Double-bridge hybrids of
MW, Broun P, Fulton TM, Giovannoni JJ, Grandillo S, Martin
Solanum bulbocastanum and cultivars of Solanum tuberosum.
GB, Messeguer R, Miller JC, Miller L, Paterson AH, Pineda
O, Roder MS, Wing RA, Wu W, Young ND (1992) High-
Johnston, SA, den Nijs TPM, Peloquin SJ, Hanneman RE Jr
density molecular linkage maps of the tomato and potato
(1980) The significance of genic balance to endosperm devel-
opment in interspecific crosses. Theor Appl Genet: 57:5–9
Thieme R, Darsow U, Gavrilenko T, Dorokhov D, Tiemann H
Lander EC, Green P, Abrahamson J, Barlow A, Daly MJ (1987)
(1997) Production of somatic hybrids between S. tuberosum L.
MAPMAKER: an interactive computer package for constructing
and late blight resistant Mexican wild potato species. Euphyt-
primary genetic linkage maps of experimental and natural pop-
Wastie RL (1991) Breeding for resistance. In: Ingram DS,
Leonards-Schippers C, Gieffers W, Salamini F, Gebhardt C (1992)
Williams PH (eds) Advances in plant pathology, vol. 7,
The R1 gene conferring race-specific resistance to Phytophth-Phytophthora infestans, the cause of late blight in potato. ora infestans in potato is located on potato chromosome V.
Young ND, Tanksley SD (1989) RFLP analysis of the size of chro-
Leonards-Schippers C, Gieffers W, Schafer-Pregl R, Ritter E,
mosomal segments retained around the TM-2 locus of tomato
Knapp SJ, Salamini F, Gebhardt C (1994) Quantitative resis-
during backcross breeding. Theor Appl Genet 77:353–359
tance to Phytophthora infestans in potato: A case study forQTL mapping in an allogamous plant species. Genetics137:67–77
DEPRESIÓN EN ADOLESCENTES Una cruz que no llega sola. (Artículo publicado en la revista “Mujer” del diario “La Tercera”, Chile) La depresión se está presentando cada vez a más temprana edad y con mayor frecuencia acompañada generalmente de otros trastornos, lo que toma por sorpresa a los padres y a la sociedad. Faltan siquiatras especializados; se abusa de los medicamen
El Juez y la Conectividad – el nuevo principio de la José Eduardo de Resende Chaves Júnior1 Aunque haya virado cliché enaltecer el potencial colectivo de la red, en el Poder Judicial eso todavía es un gran tabú, pues la decisión judicial, mismo cuando proferida de forma colegiada, es fruto todavía de un proceso de convicción profundamente monocrático. Sin perquirir sobre las compleja
 AGA TTC TTG GAC/ TGA GGT TGA TAG TGG GTG), wereused to generate fragments from both fusion parents using lowstringency polymerase chain reaction (PCR) conditions (see be-low). The resulting monomorphic fragments were cloned as de-scribed. Sequence differences between the CT88 plasmid insertsderived from the two fusion parents (PT29 and R4) were used todesign new primers specific to the resistant parent, PT29 (CT88forward and reverse: TGA GGC TCT TGG CTT TCG/AAT CAATTG AGC ATC TTG AGA).
AGA TTC TTG GAC/ TGA GGT TGA TAG TGG GTG), wereused to generate fragments from both fusion parents using lowstringency polymerase chain reaction (PCR) conditions (see be-low). The resulting monomorphic fragments were cloned as de-scribed. Sequence differences between the CT88 plasmid insertsderived from the two fusion parents (PT29 and R4) were used todesign new primers specific to the resistant parent, PT29 (CT88forward and reverse: TGA GGC TCT TGG CTT TCG/AAT CAATTG AGC ATC TTG AGA).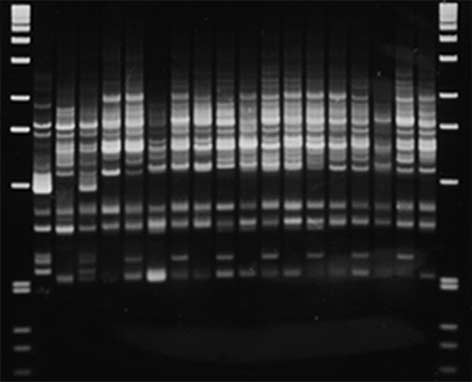 P2 SH T1 BC1 T2
P2 SH T1 BC1 T2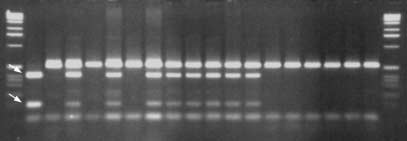
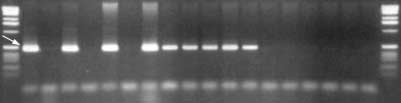 population it is possible that the fragments scored are
SCAR MARKER P09
population it is possible that the fragments scored are
SCAR MARKER P09