Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Geographic patterns of genetic variation in brushtail possums trichosurus vulpecula and implications for pest control
1 School of Biological Sciences, Victoria University, P.O. Box 600, Wellington.
Present address: Science Directorate, Department of Conservation, P.O. Box 10420, Wellington.
2 Forest Research Institute, P.O. Box 31-011, Christchurch.
Present address: Advocacy and Extension Directorate, Department of Conservation, P.O. Box 10420, Wellington.
GEOGRAPHIC PATTERNS OF GENETIC VARIATION INBRUSHTAIL POSSUMS TRICHOSURUS VULPECULA ANDIMPLICATIONS FOR PEST CONTROL
Summary: Two morphological types of brushtail possum (Trichosurus vulpecula) were introduced to New Zealand: smaller, grey possums from mainland southeastern Australia, and larger, black possums from Tasmania. Analysis of patterns of allozyme variation and allele frequencies of present-day possum populations in New Zealand and southeastern Australia indicates that populations comprised predominantly of black possums remain genetically similar to possums in Tasmania, whereas predominantly grey populations are genetically closer to Victorian and New South Wales possums. The distribution of possums in New Zealand can be accounted for at least partly by selection of stock types with respect to climate. Genetic differences between populations may have important implications for the control of possums, because Tasmanian possums have a greater resistance than mainland southeastern Australian possums to 1080 poison (sodium monotluoroacetate), which is commonly used to control possums in New Zealand. Keywords: Genetic variation; allozyme electrophoresis; brushtail possum; Trichosurus vulpecula; introduced species; pest control; selection.
IntroductionStudies of the genetics of introduced species are often
population had an overall observed heterozygosity of
limited by lack of an historical context in which to
0.029 and was fixed for the common allele at three
interpret patterns of genetic change, because the
loci which were variable in populations from New
origins, genetics, and history of introduced stock are
South Wales and Victoria. A Victorian population had
seldom known. The introduction of the brushtail
a heterozygosity of 0.040 and was fixed at two loci
possum (Trichosurus vulpecula Kerr: Marsupialia) to
variable in Tasmania and New South Wales, whereas
New Zealand, on the other hand, has been relatively
a New South Wales population had a heterozygosity
well documented, as has its morphology, distribution,
of 0.048 (Triggs, 1987). Thus the level of genetic
and ecology (Morgan and Sinclair, 1983). These
(allozymic) variation in New Zealand populations
factors provide reference points for evaluating
should depend on the degree of mixing of these
patterns of genetic variation, gene frequencies, and
Australian stocks, as well as on any changes that have
More than 200 possums were imported to New
According to historical records (Pracy, 1962),
Zealand from Australia between 1837 and 1924 in
order to establish a fur industry (Pracy, 1962). (Trichosurus vulpecula vulpecula) were small and grey,
Although the possum is commercially important for
whereas the Tasmanian stock (Trichosurus vulpecula
its fur in New Zealand (Pracy, 1981), it is also a
fuliginosa), although probably polymorphic for colour
major pest. Possums cause damage to native and
(Kean, 1971), was larger and black. In New Zealand,
exotic forests (Bathgate, 1973), erosion control
possums can be classified as either 'black' or 'grey',
plantings (Jolly and Spurr, 1981), crops (Spurr and
although a range of shades occurs from black through
Jolly, 1981), pasture (Gilmore, 1965), orchards (Anon,
brown, red-brown, and grey-brown to silver-grey.
1968), and nectar sources (Anon, 1973), as well as
Mixed populations, having both grey and black
being a reservoir for bovine tuberculosis (Ekdahl,
individuals, occur in many parts of New Zealand, but
Smith and Money,. 1970). The significance of the
the distribution of coat colours is not even in different
possum as a competitor of native birds has also been a
parts of the country (Wodzicki, 1950; Kean, 1971).
cause for concern (Leathwick, Hay and Fitzgerald,
Some areas, such as Westland, have almost all black
1983; Fitzgerald, 1984; Wardle, 1984).
possums, whereas other areas, such as Northland,
have only grey possums. Body size also varies between
from Victoria, New South Wales, and Tasmania
areas (Yom Tov, Green and Coleman, 1986; our
(Pracy, 1962). These Australian populations differ in
both the amount of heterozygosity and in the number
This mosaic of coat colours and body size may be
of variable loci (Triggs, 1987). A Tasmanian
the result of a) a non-random pattern of introduction
New Zealand Journal of Ecology 12:New Zealand Ecological Society
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12, 1989
coupled with a subsequent lack of natural dispersal
predominantly grey New Zealand populations are
over long distances, b) a haphazard pattern of
most similar to those of mainland Australian
liberations followed by selection, c) random chance,
populations. A non-random distribution of Australian
or d) most likely, a combination of processes. There is
stocks in New Zealand may have implications for pest
some evidence that predominantly one colour morph
control, as Tasmanian possums are more resistant to
was liberated in certain areas. Most possums liberated
1080 poison at low temperatures than mainland
in Westland, for example, were black (Pracy, 1962).
Australian (New South Wales) possums (McIlroy,
However, liberation records reveal that both black and
1983). The analysis is complicated by any genetic
grey possums were introduced to most areas of New
changes that have accompanied the colonization of
Zealand (Pracy, 1962), suggesting that the pattern of
New Zealand by small founder populations (Triggs,
distribution of possums in New Zealand has resulted
1987) and by the untestable assumption that the
Australian populations that we sampled accurately
The main aim of our study was to use allozyme
estimate the allele frequencies of possums originally
electrophoresis to determine whether the non-random
distribution of colour morphs in New Zealand isaccompanied by an associated pattern of allelic
distribution which can be related to the different
Samples of liver, muscle, and blood were collected
Australian stocks. If coat colour does reflect the
from possums in four locations in southeastern
origins of New Zealand populations with respect to
Australia and 10 locations on New Zealand's North,
Tasmanian and mainland Australian stocks, then two
South and Stewart Islands (Table 1). The sample from
predictions' can be made: (1) that the amount of
South Australia was collected for use as an outgroup,
variation (heterozygosity and polymorphism) in each
because possums are not known to have been exported
New Zealand population depends on the proportions
to New Zealand from South Australia. Specimens
of the two colour morphs in the population (in
were frozen on dry ice or in liquid nitrogen in the
particular, predominantly black populations should
field and stored in an ultra-cold (-80°C) freezer for
have a lower level of variation than grey populations
the duration of the study. For electrophoretic analysis
and mixed-colour populations a higher level of
small sub-samples of tissues were macerated in an
variation than non-mixed populations); (2) that allelic
equal volume of distilled water, then centrifuged at
frequencies in predominantly black New Zealand
2000 rpm for 5 minutes. The resulting supernatant
populations are most similar to those of Tasmanian
fractions were subjected to starch-gel electrophoresis,
populations, whereas allele frequencies in
Table 1: Sampling locations, sample sizes, and meteorological data (mean annual rainfall MAR and mean annual temperatureMA T). Meteorological data are from New Zealand Meteorological Service Misc. Publ. 177 (1981) or the Tasmanian YearBook (1985).*Sample collected by Ecology Division, DSIR.Australia New Zealand
TRIGGS and GREEN: GENETIC VARIATION IN POSSUMS
using gels made of 14% Electrostarch (Madison,
Wisconsin, lot no. 392) and modifications of the
methods of Selander et al. (1971), Harris and
A total of 25 enzymes and 7 general proteins
Hopkinson (1976), and Allendorf et al. (1977), as
(including haemoglobin), encoding 45 loci, was
described in Triggs (1987). The recommendations of
resolved: aconitase (Acon 1-2, E.C. no. 3.1.3.2),
Murphy and Crabtree (1985) were followed in
adenylate kinase (Ak, 2.7.4.3), B-galactosidase (B-Gal,
labelling enzymes, genetic loci, and alleles.
3.2.1.23), creatine kinase (Ck, 2.7.3.2), diaphorase
For each population, the level of genetic variation
(Dia, 1.6.2.2), erythrocyte acid phosphatase (Eap,
was assessed by degree of polymorphism (P) and
3.1.3.2), esterase (list 1-6, 3.1.1.1), general proteins
observed heterozygosity (H). Genetic differentiation
(Gp 1-6), glucose-6-phosphate dehydrogenase (Gd,
between populations was estimated using Nei's (1978)
1.1.1.49), glucose phosphate isomerase (Gpi, 5.3.1.9),
unbiased genetic distance (D); populations were then
clustered using the UPGMA algorithm (Sneath and
dehydrogenase (Glud, 1.4.1.3), glutamate oxaloacetate
Sokal, 1973). All data were analysed using the
transaminase (Got 1-2, 2.6.1.1), glycerol-3-phosphate
BIOSYS-l programme (Swofford and Selander, 1981).
dehydrogenase (Gpd, 1.1.1.8), haemoglobin (Hb),
Correlation coefficients, r, were used to determine
isocitrate dehydrogenase (Icd 1-2, 1.1.1.42), lactate
the relationships between allele frequency at each
dehydrogenase (Ldh 1-2, 1.1.1.27), malate
locus (Table 2), coat colour (given by "% black" -
dehydrogenase (Mdh 1-2, 1.1.1.37), malic enzyme
the percentage of black possums in a population;
(Me, 1.1.1.40), mannose phosphate isomerase (Mpi,
Table 3), mean adult body length and weight in each
population (Table 3), latitude (Table 1), and climate
phosphogluconate dehydrogenase (Pgd, 1.1.1.44),
(mean annual rainfall and mean annual temperature;
phosphoglucomutase (Pgm 1-2, 2.7.5.3), purine
Table 2: Allele frequencies at polymorphic loci, % polymorphic loci (P), and observed heterozygosity (H) in southeasternAustralian and New Zealand populations of Trichosurus vulpecula. (Populations numbered as in Table 1.) N = sample size.
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12, 1989
Table 3: Morphological characteristics of New Zealandpopulations of brush tail possum. Equal numbers of adult
Mean estimates of allozyme variation in New
females and adult males were sampled.
Zealand (P = 0.155, H = 0.041) were slightly, but
not significantly, greater than those of the Australian
stock populations (P = 0.141, H = 0.039; Table 4).
Comparisons of polymorphism and heterozygosityamong Australian and New Zealand populations
(Table 4) suggest that the amount of variation in New
Zealand depends to some extent on the proportion of
each colour morph in the population. New Zealand
populations with more than 50% black individuals
had a significantly lower level of variation than
predominantly grey populations (t = 2.37, p < 0.05
for P; t = 3.4, p < 0.01 for H), in parallel with the
lower level of variation in Tasmanian compared to
mainland Australian populations (Table 4). Mixed
nucleoside phosphorylase (Np, 2.4.2.1), sorbitol
colour populations in New Zealand had a higher level
dehydrogenase (Sordh, 1.1.1.14), superoxide dismutase
of variation than non-mixed populations, as expected
(Sod 1-2, 1.15.1.1), and unidentified dehydrogenase
from the mixing of stocks fixed for different loci,
although the difference in variation was not significant
Allozyme variation in New Zealand populations
between mixed and non-mixed populations (t = 2.07,
No alleles were detected in New Zealand that were not
P = 0.07 for P; t = 1.30, p = 0.2 for H). The
found in at least one Australian population (Table 2).
difference in variation between mixed New Zealand
TRIGGS and GREEN: GENETIC VARIATION IN POSSUMS
Table 4: Mean and standard deviation of polymorphism (P) and heterozygosity (H) for brush tail possum populations ofsimilar geographic origin or coat colour, and for Australian stocks (.*excludes Adelaide sample).
New Zealand mixed colour (32-66%black possums)
New Zealand non-mixed (0-2%, 73-100% black possums)
New Zealand black (53-100% black possums)
and Australian stock populations was not significant
Genetic relationships among New Zealand and
(t = 1.31, p = 0.2 for P; t = 0.4, p = 0.5 for H)
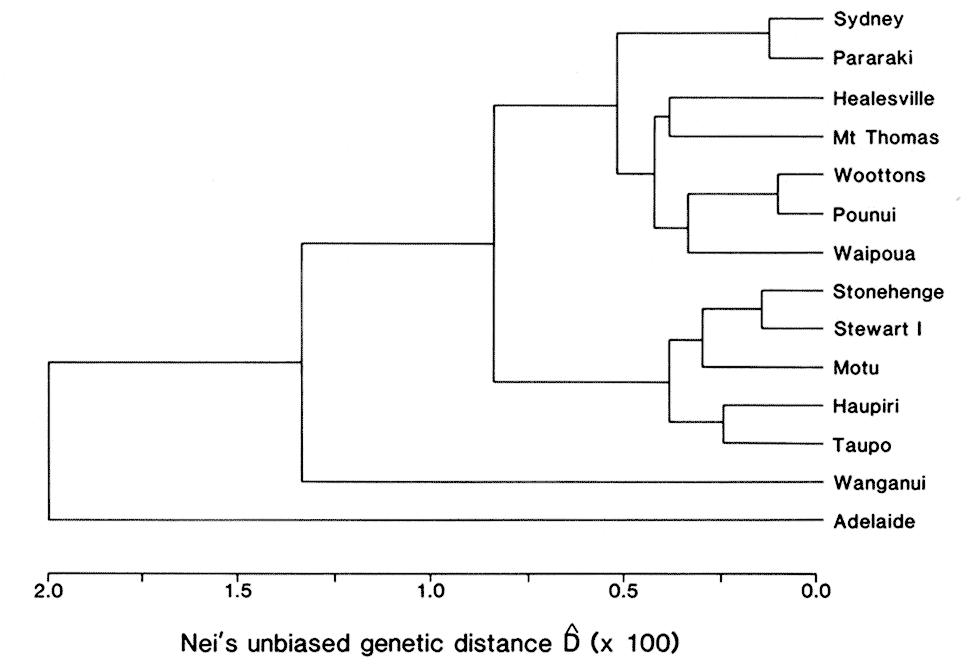
Two main clusters of populations were identified by
The level of variation was not entirely consistent
phenetic clustering, based on Nei's D (Fig. 1). New
among New Zealand populations with similar
Zealand populations with a high proportion of grey
proportions of each colour morpho For example,
possums (0-53% black) and grey Australian
Waipoua 0% black) had a 50% greater heterozygosity
populations were closely associated, as were
than Wanganui (2% black), presumably as a result of
predominantly black New Zealand populations
genetic drift in small founder populations. Changes in
(61-100% black) and the Tasmanian population. The
levels of variation associated with colonization are
inclusion of New South Wales in the former cluster
may be due to either the small genetic distance
Figure 1: UPGMA phenogram of genetic relationships among southeastern Australian and New Zealand populations ofbrushtail possum.
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12, 1989
colour and mean daily temperature (r = -0.77,
confirmation that some possums were imported from
p< 0.01), but not with latitude (r = 0.48, p > 0.05).
New South Wales. The Me-1(c) allele found in many
Several significant correlations were also found
New Zealand populations was also found in 'possums
between allele frequencies, coat colour, latitude and
from New South Wales but not in the sample from
climate in New Zealand. A total of 56 correlations
Victoria, although it is also possible that this allele
was calculated. With a 5% chance of a type II error,
was present at low frequency in Victoria. However,
only 2.8 significant correlations were expected by
the greater genetic similarity of New Zealand grey
chance, but 7 were observed. Colour (% black) was
populations to Victoria (Fig. 1) and the presence in
significantly (p < 0.05) correlated with allele
New Zealand of several alleles not found in the New
frequency for Est-1(a), Est-1(b), Icd-2(a), Icd-2(c),
South Wales sample [Est-1(c), Idh-2(c) and Me-1(b)]
and Pgd-1(a). Mean annual temperature was
suggest the predominance of Victorian stock in New
significantly correlated with Pgd-1(a) (p < 0.05) and
associated with Est-1(b), Est-5(a) and Got-1(a)
(p < 0.10). Latitude was correlated with Pgd-1(a)
resemble those from New Zealand. This result was
(p < 0.05). No significant correlations were found
expected, because no possums were imported to New
between allele frequency and rainfall. Colour and
Zealand from South Australia. One population,
allele frequency appear to be the most closely
Wanganui, did not fall into either cluster, possibly due
associated; selection of one or both may be linked to
to genetic drift, if the Wanganui population had a
temperature. No significant correlation (r = 0.012,
P > 0.05) was found between overall heterozygosity
Although the genetic distances involved are very
and latitude, in contrast to the result for Australia
small (the 'black/grey' separation occurs at D =
0.008), our analysis suggests that the origins of New
Body length and weight were also correlated with
Zealand populations are still reflected in their allele
coat colour in our study populations (r = 0.68 and
frequencies and may be roughly estimated by coat
r = 0.64 respectively; p < 0.05). Yom Tov (1984) and
colour. The relationship between coat colour in New
Yom Tov et al. (1986), in an extensive morphological
Zealand populations and position within the genetic
survey of possums in New Zealand, found significant
cluster is by no means perfect. Presumably random
negative correlations between many skull and body
genetic drift in the relatively small founder
length measurements and mean annual temperature.
populations has led to changes in gene frequency in
However, they did not consider coat colour, and our
many populations. However, the general pattern that
reanalysis of data on body and skull measurements in
emerges, grouping predominantly black populations
Yom Tov (1984) with respect to coat colour produced
with the Tasmanian sample and grey with mainland
better correlations between body size and % black
Australia, is unlikely to have arisen by chance (that is,
than between body size and mean annual temperature
by random drift). The implication is that either
for all characters except distance between bullae
interbreeding of stock types for 150 years has been
insufficient to establish a panmictic unit, or that
Table 5: Correlations (r) between body and skull
selection has acted differentially on stock types to
measurements, coat colour (% Black), and mean annual
produce genetic structuring of allozymes and colour
temperature (MA T). Body and skull measurements andcorrelation coefficients of measurements vs MAT are from
Selection as an explanation for the mosaic of
Yom Tov (/984). % Black (the proportion of black possums
colour morph distribution in New Zealand is
in a population) values are from our data. Significance levels
supported by evidence of correlations between coat
of r: .*p < 0.05, .**p < 0.001.
colour (% black possums in a population), allelefrequencies, body size and weight, and climatic
variables. A significant correlation (r = 0.642, p <
0.01) exists between coat colour and rainfall in New
Zealand (R.E. Brockie, pers. comm.). Data from our
10 mainland New Zealand study areas showed a
similar, although non-significant, correlation between
colour (% black) and rainfall (r = 0.58, P < 0.10). A
significant correlation in our data was found between
TRIGGS and GREEN: GENETIC VARIATION IN POSSUMS
sparrows in North America (Johnson and Selander,
The introduction of possums from at least two regions
1964). In all cases, these associations have developed
of Australia, followed by hundreds of largely
very rapidly, within a few hundred years, suggesting
undocumented liberations of New Zealand-bred stock,
that selection for local adaptation may be very strong,
has produced a complex pattern of genetic
even in small populations in which random forces are
relationships in New Zealand possums, upon which
selective and random genetic changes have been
Our results have important implications for the
control of possums in New Zealand. Currently, large-
Our results generally conform to the prediction
scale control of possum numbers is by aerially-sown
that if coat colour of New Zealand populations
cereal baits or carrots with 1080 (sodium
indicates origin, then (1) the level of genetic variation
monofluoroacetate) poison. One serious concern is the
in New Zealand populations should be related to the
dosage level of 1080 required to kill a possum. Bell
proportion of each colour morph in a population, and
(1972), Rammell and Fleming (1978), and McIlroy
(2) allele frequencies of New Zealand populations with
(1983) experimentally determined an LD of about 0.8
different proportions of each coat colour should
mg 1080 kg-1 body weight for possums. In contrast,
reflect allele frequencies of the Australian stock types.
the New Zealand Forest Service found an LD of
Thus, mainland Australian and Tasmanian stocks are
1.3-2.1 mg kg-1 (Anon, 1978), requiring a toxic
not distributed at random in New Zealand. Black and
loading of 0.15% w/w on baits of mean weight of 4
grey populations also differ in average body size (Yom
g. The higher dose not only adds to the cost of
Tov et al., 1987; our study), as they do in Australia
poisoning operations and the risk to non-target
species, but also leads to a high aversion rate as some
Selection with respect to climate appears to be an
possums can detect and reject 1080 at concentrations
important determinant of the distribution of possum
of 0.1 % w/w or more (Morgan, 1982). At present,
types in New Zealand, although deliberate
flavours such as cinnamon are used as masks to
introductions of possums of different stocks to
disguise the poison (Morgan, Batcheler and Peters,
suitable habitats probably also played a part in
1986), and baits are loaded either at 0.08% w/w or
determining present distributions. In New Zealand,
0.15% w/w (D.R. Morgan, pers. comm.) as a result
cold, wet areas tend to harbour large, black possums
of the ambiguous data published on the possum's
most similar to the Tasmanian type, whereas warm,
sensitivity to 1080 poison. The cause of differences in
dry areas harbour small, grey, mainland Australian-
LD between laboratories is still not clear, although
type possums. Areas of intermediate climate have
variation in the techniques for handling 'and
mixed populations. The significant correlations
acclimatising possums were probably partly
between temperature and allele frequency, colour and
responsible (Anon, 1979). McIlroy (1983) found
body size of possums in New Zealand suggest that
neither acclimatisation nor stress had any effect on the
these were either directly selected for or acted as
LD but he did find significant differences between
markers for other characteristics selected as possums
possums from different regions of southeastern
colonized New Zealand. The correlation between
Australia. At low temperature (10°C) Tasmanian
colour and rainfall has been found even within a
possums were more resistant to 1080 than mainland
single valley with a steep rainfall gradient (R.E.
(New South Wales) possums. The LD,. for Tasmanian
Brockie, pers. comm.), emphasizing the strength of
possums was 0.92 mg kg-1, while New South Wales
selection. An association between coat colour and
possums had an LD of 0.42 mg kg-1, (McIlroy,
rainfall has also been documented in Tasmania
1983). Decreased sensitivity to 1080 at low
(Guiler, 1953), black possums being more common in
temperatures has also been found in racoons (Eastland
areas of high rainfall. A physiological basis for
and Beasom, 1986). Much higher tolerances to 1080
different climatic tolerances is suggested by the
occur in brushtail possums and other mammals in
differences in water metabolism between black and
Western Australia, where high levels of fluoroacetates
grey morphs (Williams and Turnbull, 1983).
occur naturally in some plants (King, Oliver and
Other studies of introduced species have also
documented associations between colour or size and
The possums used in the original New Zealand
climate. These studies include house sparrows (Baker,
Forest Service trials (Anon, 1978) were from an area
1980), mynas (Baker and Moeed, 1979), and stoats
of predominantly black possums, and were therefore
(King and Moody, 1982) in New Zealand, and house
likely to be of the Tasmanian type, whereas the
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12,1989
possums used in the Ministry of Agriculture and
P.E. Cowan, C.H. Daugherty, P.J. Moors, R.M.
Fisheries trials (Bell, 1972) were from an area of grey
Sadleir, P.J. Smith, and D.R. Towns for their advice
possums and were therefore likely to be of the
mainland Australian type. A higher tolerance wouldtherefore be expected in the New Zealand Forest
Service results. However, a reciprocal exchange ofpossums between the two testing laboratories still
Allendorf, F.W.; Mitchell, N.; Ryman, N.; Stahl, G.
resulted in different LD estimates (Anon, 1979),
1977. Isozyme loci in brown trout (Salmo trulla
suggesting that some other factor, such as handling
L): Detection and interpretation from population
technique, was also involved. The temperature regime
used in each laboratory was not given and could have
Anon. 1968. Oppossum destruction. Orchardist of
If Tasmanian possums are more resistant to 1080,
Anon. 1973. Damage to nectar sources by opossum
as found by McIlroy (1983), then there is a good case
and deer. New Zealand Beekeeper 35: 75.
for using a higher dose rate of 1080 in cold, wet areas,
Anon. 1978. Bait development and toxicology. New
where Tasmanian-type possums predominate, than in
Zealand Forest Service, Forest Research Institute
warm, dry areas, where mainland-type possums
predominate. Tasmanian possums are also heavier and
Anon. 1979. Bait development and toxicology. New
larger (Yom Tov and Nix, 1986; Triggs, 1987,
Zealand Forest Service, Forest Research Institute
Appendix II). In order to receive a lethal dose they
would need to consume more baits at the same toxic
Baker, A.J. 1980. Morphometric differentiation in
loading than would smaller, lighter, mainland-type
New Zealand populations of the house sparrow
possums. As a hypothetical example, assuming that a
(Passer domesticus). Evolution 24: 638-653.
totally grey population has an equivalent LD and
Baker, A.J.; Moeed, A. 1979. Evolution in the
mean body weight to mainland Australian possums
introduced New Zealand populations of the
(i.e. 0.42 mg 1080 kg-1 and 2.3 kg) and a black
common myna, Acridotheres tristis (Aves:
population is equivalent to Tasmanian possums (0.92
Sturnidae). Canadian Journal Zoology 57:
mg 1080 kg-1 and 3.1 kg), then 50070 of the grey
population would be killed with 0.97 mg 1080 per
Bathgate, J.L 1973. Summary of questionnaire
possum, whereas the black population would require
returns. In: Assessment and management of
an average of 2.9 mg 1080 per possum. introduced animals in New Zealand forests, pp. 102-116. New Zealand Forest Service, Forest
Research Institute Symposium No. 14.
Bell, J. 1972. The acute toxicity of four common
This research was supported by a Postgraduate Grant
poisons to the opposum, Trichosurus vulpecula.
to S. Triggs from Ecology Division, Department of
New Zealand Veterinary Journal 20: 212-214.
Scientific and Industrial Research. Technical support
Eastland, W.G.; Beasom, S.L 1986. Effects of
and additional finance were provided by Victoria
University of Wellington. For assistance with sample
racoons. Wildlife SocietyBulletin 14: 234-235.
collections we are indebted to staff of Forest Research
Ekdahl, M.a.; Smith, B.L.; Money, B.L. 1970.
Institute, Christchurch; Ecology Division, DSIR;
Tuberculosis in some wild and feral animals in
Rotorua-Taupo, Western, and Wairarapa Pest
New Zealand. New Zealand Veterinary Journal
Destruction Boards; Wildlife Service; David Crockett
of Taiko Camp; staff of Macquarie University,
Fitzgerald, A.E. 1984. Diet overlap between kokako
University of Adelaide, Victoria Fisheries and Wildlife
and the common brushtail possum in central
Service, Monash University, and Tasmanian National
North Island, New Zealand. In: Smith, A.P. and
Parks and Wildlife Service; and the Dunbabbin
Hulme, J.D. (Editors), Possums and gliders, pp.
family. Permits for collecting, exporting and
569-573. Australian Mammal Society, Sydney.
importing possum tissues were supplied by the
Australian National Parks and Wildlife Service of
Gilmore, D.P. 1965. Opposums eat pasture. New
New South Wales, South Australia, Victoria and
Zealand Journal of Agriculture 110: 284-286.
Tasmania, and the Ministry of Agriculture and
Guiler, E.R. 1953. Distribution of the brush possum
Fisheries, New Zealand. We thank Drs R.E. Brockie,
in Tasmania. Nature 172: 1091-1093.
TRIGGS and GREEN: GENETIC VARIATION IN POSSUMS
rattlesnake, Crotalus viridis viridis. Herpetologia
Harris, H.; Hopkinson, D.A. 1976. Handbook ofenzyme electrophoresis in human genetics. North-
Nei, M. 1978. Estimation of average heterozygosity
and genetic distance from a small number of
Johnson, R.F.; Selander, R.F. 1964. House sparrows:
individuals. Genetics 89: 583-590.
rapid evolution of races in North America.
New Zealand Meteorological Service. 1981. Summariesof climatological observations to 1980. New
Jolly, J.N.; Spurr, E.B. 1981. Damage by possums
Zealand Meteorological Service Miscellaneous
(Trichosurus vulpecula) to erosion-control
plantings. In: Bell, B.D. (Editor), Proceedings ofthe First Symposium on Marsupials in New
Pracy, LT. 1962. Introduction and liberation of theZealand, pp. 205-210. Zoology Publication No. opossum (Trichosurus vulpecula) into New
74, Victoria University, Wellington. Zealand. New Zealand Forest Service Information
Kean, R.I. 1971. Selection for melanism and low
Series No. 45, New Zealand Forest Service.
reproductive rate in Trichosurus vulpecula
Pracy, LT. 1981. Opossum survey. Counterpest 5:
(Marsupialia). Proceedings of the New ZealandEcological Society 18: 42-47.
Rammell, E.G.; Fleming, P.A. 1978. Compound
King, C.M.; Moody, J.E. 1982. The biology of the
1080. Properties and use of sodium
stoat (Mustela erminea) in the National Parks of
monofluoroacetate in New Zealand. Animal
New Zealand. III. Morphometric variation in
Health Division, Ministry of Agriculture and
relation to growth, geographical distribution, and
colonisation. New Zealand Journal of Zoology 9:
Selander, R.K.; Smith, M.H.; Yang, S.Y.; Johnson,
King, D.R.; Oliver, A.J.; Mead, R.J. 1978. The
polymorphisms and systematics in the genus
adaptation of some Western Australian mammals
Peromyscus. I. Variation in the old-field mouse,
to food plants containing fluoroacetate. Peromyscus polionotus. Studies in genetics,Australian Journal of Zoology 26: 699-712. University of Texas Publication 7103: 49-90.
Leathwick, J.R.; Hay, J.R.; Fitzgerald, A.E. 1983.
Sneath, P.H.A.; Sokal, R.R. 1973. Numericaltaxonomy. W.H. Freeman, San Francisco. 359 p.
mammals on the decline of the North Island
kokako. New Zealand Journal of Ecology 6:
possumsTrichosurus vulpecula to farm crops and
pasture.In: Bell, B.D. (Editor), Proceedings of the
McIlroy, J.C. 1983. The sensitivity of the brushtail
FirstSymposium on Marsupials in New Zealand, pp.
possum (Trichosurus vulpecula) to 1080 poison.
197-203. Zoology Publication No. 74, Victoria
New Zealand Journal of Ecology 6: 125-131.
Morgan, D.R. 1982. Field acceptance of non-toxic and
Swofford, D.L; Selander, R.B. 1981. BIOSYS-I:
toxic baits by populations of the brushtail possum
(Trichosurus vulpecula Kerr). New Zealand
analysis of electrophoretic data in population
genetics and systematics. Journal of Heredity 72:
Morgan, D.R.; Batcheler, C.L; Peters, J.A. 1986.
Tasmanian Year Book. 1985. Australian Bureau of
operations? In: Salmon, J.P. (Editor),
Proceedings 12th Vertebrate Pest Conference, pp.
Triggs, S.J. 1987 (unpublished). Population and
210-214. University of California, Davis,
ecological genetics of the brush-tailed possum
Trichosurus vulpecula in New Zealand. PhD
Morgan, D.R.; Sinclair, M.J. 1983. A bibliography of
Thesis, Victoria University of Wellington. 146 p. the brush-tailed possum (Trichosurus vulpecula
Wardle, J. 1984. The New Zealand beeches. Ecology,Kerr). Forest Research Institute Bulletin No. 25,
utilisation and management. Caxton Press,
Protection Forestry Division, Forest Research
Williams, C.K.; Turnbull, H.L 1983. Variations in
Murphy, R.W.; Crabtree, C.B. 1985. Evolutionary
seasonal nutrition, thermoregulation and water
aspects of isozyme patterns, number of loci, and
balance in two New Zealand populations of the
tissue-specific gene expression in the prairie
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12, 1989
common, brush-tail possum, Trichosurusvulpecula (Phalangeridae). Australian Journal ofZoology 31: 333-343.
Wodzicki, K.A. 1950. Introduced mammals of New
Zealand (an ecological and economic survey). DSIR Bulletin 98: 19-49.
Yom Tov, Y. 1984 (unpublished). Morphologicaltrends in the brush tail possum, Trichosurusvulpecula, in New Zealand. Report to EcologyDivision, Department of Scientific and IndustrialResearch, Lower Hutt. 18 p.
Yom Tov, T.; Green, W.Q.; Coleman, J.D. 1986.
Morphological trends in the common brushtailpossum, Trichosurus vulpecula, in New Zealand. Journal of Zoology 208: 583-593.
Yom Tov, Y.; Nix, H. 1986. Climatological correlates
for body size of five species of Australianmammals. Biological Journal of the LinnaeanSociety 29: 245-262.
GBS (Group B Streptococcus) First, you should know that Group B Strep is a bacterium that lives in the intestines and reproductive areas of about a third of the healthy population. It is normally not something to think about. The reason the subject comes up in pregnancy is that GBS is more dangerous to certain groups of people, including infants, than to the general population. When Mom car
 TRIGGS and GREEN: GENETIC VARIATION IN POSSUMS
Table 4: Mean and standard deviation of polymorphism (P) and heterozygosity (H) for brush tail possum populations ofsimilar geographic origin or coat colour, and for Australian stocks (.*excludes Adelaide sample).
New Zealand mixed colour (32-66% black possums)
New Zealand non-mixed (0-2%, 73-100% black possums)
New Zealand black (53-100% black possums)
and Australian stock populations was not significant
Genetic relationships among New Zealand and
(t = 1.31, p = 0.2 for P; t = 0.4, p = 0.5 for H)
Two main clusters of populations were identified by
The level of variation was not entirely consistent
phenetic clustering, based on Nei's D (Fig. 1). New
among New Zealand populations with similar
Zealand populations with a high proportion of grey
proportions of each colour morpho For example,
possums (0-53% black) and grey Australian
Waipoua 0% black) had a 50% greater heterozygosity
populations were closely associated, as were
than Wanganui (2% black), presumably as a result of
predominantly black New Zealand populations
genetic drift in small founder populations. Changes in
(61-100% black) and the Tasmanian population. The
levels of variation associated with colonization are
inclusion of New South Wales in the former cluster
may be due to either the small genetic distance
Figure 1: UPGMA phenogram of genetic relationships among southeastern Australian and New Zealand populations ofbrushtail possum.
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12, 1989
colour and mean daily temperature (r = -0.77,
confirmation that some possums were imported from
p< 0.01), but not with latitude (r = 0.48, p > 0.05).
TRIGGS and GREEN: GENETIC VARIATION IN POSSUMS
Table 4: Mean and standard deviation of polymorphism (P) and heterozygosity (H) for brush tail possum populations ofsimilar geographic origin or coat colour, and for Australian stocks (.*excludes Adelaide sample).
New Zealand mixed colour (32-66% black possums)
New Zealand non-mixed (0-2%, 73-100% black possums)
New Zealand black (53-100% black possums)
and Australian stock populations was not significant
Genetic relationships among New Zealand and
(t = 1.31, p = 0.2 for P; t = 0.4, p = 0.5 for H)
Two main clusters of populations were identified by
The level of variation was not entirely consistent
phenetic clustering, based on Nei's D (Fig. 1). New
among New Zealand populations with similar
Zealand populations with a high proportion of grey
proportions of each colour morpho For example,
possums (0-53% black) and grey Australian
Waipoua 0% black) had a 50% greater heterozygosity
populations were closely associated, as were
than Wanganui (2% black), presumably as a result of
predominantly black New Zealand populations
genetic drift in small founder populations. Changes in
(61-100% black) and the Tasmanian population. The
levels of variation associated with colonization are
inclusion of New South Wales in the former cluster
may be due to either the small genetic distance
Figure 1: UPGMA phenogram of genetic relationships among southeastern Australian and New Zealand populations ofbrushtail possum.
NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 12, 1989
colour and mean daily temperature (r = -0.77,
confirmation that some possums were imported from
p< 0.01), but not with latitude (r = 0.48, p > 0.05).