Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Effects of the antibiotic ciprofloxacin on stream microbial communities and detritivorous macroinvertebrates
Environmental Toxicology and Chemistry, Vol. 25, No. 6, pp. 1598–1606, 2006
EFFECTS OF THE ANTIBIOTIC CIPROFLOXACIN ON STREAM MICROBIAL
COMMUNITIES AND DETRITIVOROUS MACROINVERTEBRATES
JONATHAN D. MAUL,† LANCE J. SCHULER,† JASON B. BELDEN,† MATT R. WHILES,‡ and MICHAEL J. LYDY*†
†Fisheries and Illinois Aquaculture Center and Department of Zoology, ‡Department of Zoology,
Southern Illinois University, Carbondale, Illinois 62901, USA
(Received 25 July 2005; Accepted 10 November 2005)
Abstract—Microbial communities play an important role in stream ecosystem processes, such as breakdown of senescent leaf litter, and as a primary nutritional source for detritivorous macroinvertebrates. Antibiotics may affect stream microbial communities and associated ecosystem processes, especially because recent stream and river monitoring programs have indicated the presence of antibiotics downstream of wastewater treatment plants. In the current study, effects of chronic exposure to the fluoroquinolone antibiotic ciprofloxacin (Cipro) were examined on stream microbial community-level physiological profiles and growth indices of detritivorous amphipods (Gammarus spp.) and caddisflies (Lepidostoma liba). Microcosm experiments were conducted using stream sediments and water, senesced leaf material (Acer saccharum), and macroinvertebrates. A shift in function of leaf-associated microbial communities (based on carbon source utilization) was observed for samples exposed to 100 g/L of Cipro for 12 d compared to control and treatments exposed to 1 and 10 g/L of Cipro. This was attributable to carbohydrate substrates, which had 2.7- to 3.5- fold lower microbial respiration than the lower concentrations and control ( p Ͻ 0.001). For detritivores, Gammarus spp. condition index did not differ among control, 0.1, and 1.0 g/L treatments after 30-d exposures ( p Ͼ 0.05). Similarly, L. liba growth rate did not vary among control, 10, and 100 g/L treatments after 45-d exposures ( p Ͼ 0.05). These results suggest that Cipro may affect leaf-associated microbial communities, but at concentrations four orders of magnitude above those detected in streams. However, effects of the antibiotic on growth and condition of detritivores were not observed. Future work should focus on identifying specific changes in stream microbial communities as a result of Cipro exposure and impacts on other aquatic species. Keywords—Antibiotic
Microbial community-level physiological profiles
INTRODUCTION
relationship occurs in the gut of many detritivores that facil-itates acquisition of resources from ingested material via mi-
Antibiotics are used extensively for human and animal
crobial processing that is not possible from endogenously de-
health applications. Because many antibiotics are not com-
pletely metabolized, they are frequently detected in effluents
Leaf litter processing by microbes and detritivores is im-
and downstream of wastewater treatment plants and animal
portant for cycling of organic carbon in stream ecosystems,
production facilities [1,2]. Antibiotics in the environment pose
because this material often represents the major energy source
a potential threat to communities and associated ecosystem
for stream food webs [9,13]. In some stream systems, Gam-
processes because of the important role bacteria have in these
marus spp. were found to process as much as 16% of the total
leaf litter input [14]. Potential impacts of reduced microbial
Ciprofloxacin (Cipro), a second-generation fluoroquinolone
activity to aquatic macroinvertebrate functional groups that
antibiotic, has been detected in stream water in Europe [1] and
feed on detritus, such as shredders and collector-gatherers,
the United States [2] at concentrations ranging from 0.01 to
would likely influence the entire stream community and mul-
0.03 g/L, and its acute toxicity on some aquatic organisms
tiple trophic levels [13]. Furthermore, decreases in litter pro-
has been reported [3]. Cipro has antimicrobial activity against
cessing in a given stream reach may influence downstream
genera commonly associated with stream water and leaf litter,
such as Achromobacter, Acinetobacter, Flavobacterium, and
The objectives of the current study were to examine the
Pseudomonas sp. [4–6]. Of the gut bacteria identified in sev-
effects of Cipro on the function of aerobic bacterial commu-
eral aquatic invertebrates [7], genera such as Aeromonas, Fla-
nities associated with leaf litter by the use of community-level
vobacterium, Pseudomonas, and Vibrio also are affected ad-
physiological profiles and to examine the effects of Cipro on
the body condition, growth, and leaf processing of detritivo-
Although fungi often are a primary food resource for stream
rous amphipods (Gammarus spp.) and caddisflies (Lepidos-
detritivores, colonization of leaf litter by bacteria facilitates
toma liba) at the microcosm scale. We predicted that Cipro
breakdown and also may improve the nutritional quality of
would alter functional attributes of the bacterial community,
this material [8,9]. Cummins [10] suggested that many detri-
affect nutrient resources, and reduce indices of detritivore
tivorous invertebrates directly consume the bacteria found on
leaf litter and other substrates, and other investigators haveshown that some stream invertebrates may obtain 20 to 40%
MATERIALS AND METHODS
of their carbon from bacteria [11]. Furthermore, a symbiotic
Sediment and water for all experiments were collected from
* To whom correspondence may be addressed (mlydy@siu.edu).
a 100-m reach of Big Creek in Union County, Illinois, USA,
Effects of ciprofloxacin on stream microcosms
and transported to the laboratory. Big Creek is a third-order
nm. Absorbance readings were taken every 8 h during the 72-
stream in the Shawnee Hills Natural Division in southern Il-
h incubation period. Readings at the 24-h incubation time point
linois. The water for Gammarus spp. experiments was col-
for each plate were used, because the time series indicated a
lected in December and January with the following average
linear phase of microbial growth (i.e., increasing respiration)
water-chemistry values: Temperature, 10.0ЊC; dissolved oxy-
at this time. Data were standardized by the average well color
gen, 11.8 mg/L; pH 7.2; and conductivity, 363.8 S/cm. Gam-
development (AWCD) method [18]. Specifically, well data
marus spp. were collected in December with dipnets at two
were divided by AWCD to normalize for inoculum density;
locations within the study reach and transported in a 19-L
normalization using this approach resolves community pat-
bucket filled with stream water to the laboratory within 2 h
terning and minimizes the influence of growth rates [18].
of collection. Water for the L. liba experiment was collected
To gain an understanding of the specific impacts potentially
in late January and early March with the following average
driving patterns in microbial communities, carbon substrate
water-chemistry values: Temperature, 14.1ЊC; dissolved oxy-
utilization was further evaluated based on the type of carbon
gen, 10.4 mg/L; pH 8.0; and conductivity, 392.0 S/cm. Lep-
source (i.e., carbohydrate, polymer, carboxylic acid, and amino
idostoma liba were collected from first-order spring seep trib-
acid) following categories outlined by Mills and Garland [18].
utaries of Big Creek located from 0.8 to 1.5 km upstream of
Optical density values normalized for AWCD were used as a
the study reach where water, sediment, and amphipods were
relative measure of microbial respiration. Data examined were
collected. They were transported to the laboratory in 3-L plas-
leaf disk–associated microbial communities after 12 d of ex-
tic bags filled with leaf litter and spring water within 4 h of
posure to Cipro and experimental conditions.
collection. Senesced, freshly abscised sugar maple (Acer sac-charum) leaves were collected by placing a tarp under pre-
Detritivore growth and leaf processing
viously identified and marked trees on the east bank of Big
Microcosms consisted of 300-ml experimental chambers
Creek within the study reach. In the laboratory, leaves were
containing approximately 50 g of sediment and 250 ml of
air-dried and then stored in plastic bags until use. Sugar maple
water. A 30-d static renewal design was selected to simulate
represents a more labile species, with low lignin and tannin
potential exposure scenarios downstream of sewage treatment
levels and higher palatability for gammarids [14], and it was
plants, where continuous antibiotic concentrations typically
the dominant tree taxa within the study reach.
occur in effluents. Approximately 170 ml of exposure waterwas replaced daily over the time course of the experiment. Microbial community-level physiological profiles
Experiments were conducted in an environmental chamber at
In the current study, shifts in function of aerobic carbon
10ЊC and a 10:14-h light:dark photoperiod to simulate envi-
using microbes (i.e., community-level physiological profiles)
ronmental conditions occurring at the stream during the period
were examined using 96-well EcoPlates (BIOLOG, Hayward,
that Gammarus spp. were collected. The effect of Cipro on
CA, USA). Changes in community-level physiological profiles
Gammarus spp. body condition and leaf processing was ex-
have been used previously for assessing changes in microbial
amined at nominal aqueous concentrations of 0.1 and 1.0 g/
communities because of environmental conditions or impacts
L. Control microcosms were identical except that 170 ml of
in aquatic environments [17]. EcoPlates consisted of triplicate
stream water without Cipro was added daily. Twelve, 9, and
wells of 31 preloaded carbon sources and were used to classify
11 replicate chambers were used for the control, 0.1, and 1.0
microbial functional groups (i.e., microbial groups that use
different carbon sources) based on respiration products reduc-
After field collection, Gammarus spp. were acclimated for
5 d to laboratory conditions in approximately 19 L of stream
Microcosms consisted of 300-ml experimental chambers
water at 10ЊC and a density of approximately 50 individuals/
containing approximately 50 g of sediment and 250 ml of
L. Maple leaves were supplied to the acclimation tank. Before
water. A static renewal design was used, with approximately
the experiment, a similarly sized cohort was collected between
170 ml of exposure water replaced daily over the time course
0.825- and 1.400-mm sieves. Individuals from this group were
of the experiment. Three A. saccharum leaf disks (diameter,
randomly distributed to experimental chambers (n ϭ 5 per
2.5 cm) were placed in each experimental chamber 5 d before
chamber), and an additional group (n ϭ 48) was immediately
the experiment, and 170 ml of stream water were renewed
preserved in 10% formalin for initial mass and length deter-
daily. After this 5-d period, one leaf disk was removed from
each experimental chamber for microbial analyses, and Cipro
Leaf disks were added to experimental chambers with Gam-
treatment began at three nominal Cipro concentrations (1.0,
marus spp. Leaf disks were prepared by rinsing A. saccharum
10, and 100.0 g/L). Experiments were conducted at 14ЊC and
leaves with deionized water to remove debris and cutting disks
a 10:14-h light:dark photoperiod, and three replicate chambers
(diameter, 1.1 cm) with a cork borer, avoiding main and sec-
were used for the control and each treatment. Additional leaf
ondary veins. Leaf disks were dried for approximately 3 d at
disks were removed from each experimental chamber at 4 and
60ЊC to constant mass. Dry initial leaf disk mass was measured
to the nearest 0.001 mg, and leaf disks were separated into
After the predetermined exposure period (0, 4, and 12 d),
five groups of 4 to 8 mg each. One disk from each group was
leaf disks were removed from chambers; placed in vials con-
strung on monofilament fishing line in random order to min-
taining 18.0 ml of sterile, deionized water; and vortexed twice
imize the variation of overall leaf mass among experimental
for 10 s to dislodge leaf-associated microbes. Sample aliquots
chambers. Thus, each experimental chamber contained a
of 100 l were inoculated into each well of the EcoPlate under
closed loop of monofilament line containing five leaf disks
a laminar flow hood. An EcoPlate was used for each leaf disk.
that were submerged and positioned on top and in contact with
Plates were covered with lids and incubated at 29.0ЊC. Ab-
the sediment. Leaf disks were preconditioned in stream water
sorbance was measured with a microtiter spectrophotometer
in experimental chambers 4 d before Cipro exposures.
(Thermo Labsystems, Vantaa, Finland) at a wavelength of 590
Gammarus spp. from the initial group and those collected
at the end of the experiment were measured for total length
for Lepidostoma spp. (0.0079 and 2.649, respectively) [21].
from digital pictures with ImageJ software (v1.32j; National
Growth rate was estimated using the instantaneous growth
rsb.info.nih.gov/ij/). All Gammarus spp. were dried at 60ЊC
G ϭ [ln (m /m )]/d
for 3 d to constant mass, and mass was estimated to the nearest
0.001 mg with a microbalance (Cahn C-33; Cahn Instruments,
where G ϭ daily instantaneous growth rate, m ϭ final dry
Cerritos, CA, USA). Organisms were ashed at 550ЊC to de-
mass, m ϭ initial dry mass, and d ϭ 45 (length of experiment
termine ash-free dry mass (AFDM). A body condition index
(BCI) was calculated for Gammarus spp. by regressing AFDMof control organisms against total body length using standard
Water chemistry and Cipro stability in test systems
major axis regression, a model II regression method [19,20].
Temperature, dissolved oxygen, conductivity, and pH were
The residuals (x ϩ 1) of all data using the control regression
measured in experimental chambers periodically throughout
equation were used as a BCI [19]; thus, a BCI value of one
the experiments using a YSI 55 water-quality meter (Yellow
indicated an AFDM that was expected for the corresponding
Springs Instrument Company, Yellow Springs, OH, USA) and
length based on the linear-regression model. In organisms with
an Oakton conductivity meter (Oakton Instruments, Vernon
a rigid exoskeleton, such as Gammarus spp., suboptimal re-
source conditions might not be reflected in length measure-
Cipro exposure solutions were prepared daily in water col-
ments alone; thus, a condition index was used that was based
lected from Big Creek. Stock Cipro solutions were prepared
on the mass expected for a particular length.
in 0.01% acetic acid in deionized water, and each stock was
At the end of the experiment, leaf disks were removed,
used for no more than 3 d. Acetic acid concentration in the
dried at 60ЊC for 3 d, and cooled in a dessicator for 24 h before
treatment chambers, as a percentage, never exceeded 5.7 ϫ
final mass determinations. Leaf disks were ashed at 550ЊC for
10Ϫ6, and this amount had no effect on pH in the naturally
35 min to determine AFDM. Leaf breakdown was estimated
buffered water system used. Cipro was obtained from Sero-
by percentage loss of AFDM after 30-d exposure to experi-
logicals Proteins (ciprofloxacin hydrochloride; Kankakee, IL,
mental conditions. To determine the AFDM for initial leaf
disks, dry and ash masses were determined for a subset of
The concentration of Cipro was determined for water sam-
initial leaf disks (n ϭ 16), and that ratio was used to convert
ples collected from the test chambers before and after renewal
measured initial dry masses to AFDM.
to provide a measure of Cipro stability within the test system.
Microcosm setup and conditions for the L. liba experiment
Samples were collected five times during the studies for the
were identical to that described above for Gammarus spp. with
higher-level Cipro treatments (10 and 100 g/L). Measure-
the following exceptions: A 45-d static renewal design was
ments of the lower-level concentrations were not conducted
used, experiments were conducted at 14ЊC, and nominal Cipro
because of poor quantitative capabilities as a result of matrix
exposure concentrations were 10 and 100 g/L, with 18 rep-
interferences at low concentrations. Because the exposure test
licate chambers for each of the control, 10, and 100 g/L
systems were nearly identical throughout the studies reported
treatments. These modifications were made because effects of
in this manuscript, data collected among individual experi-
Cipro on body condition and leaf processing by Gammarus
ments were pooled to provide a single measure of stability.
spp. were not observed at the lower concentrations, and 14ЊC
During each sample collection in the detritivore experi-
was a more appropriate temperature for optimal L. liba growth.
ments, 10 ml of water were withdrawn from four replicate
After collection from spring seeps, L. liba were held in the
samples in the same treatment and pooled together to make
laboratory in stream water with A. saccharum leaves for 3 d
40 ml. For the microbial experiment, a 40-ml composite sample
before use in experiments. Initial total length of randomly
was derived from 13, 13, and 14 ml collected from each of
selected individuals was measured to the nearest 0.5 mm using
the three experimental units for each treatment. Samples were
a dissecting microscope, and three L. liba were distributed to
adjusted to 5% NH OH and immediately extracted using solid-
each experimental chamber (mean, 3.4 mm; range, 2.5–5.0
phase extraction (SPE). The SPE (MAX, 60 mg; Waters, Mil-
mm). Total length measurements were obtained by gently
ford, MA, USA) was preconditioned with 1 ml of methanol
scraping off the terminal end of the portable larval case, ex-
followed by 1 ml of 5% NH OH in deionized water. The sample
posing both the head and terminal end of the organism. Or-
was then eluted at a rate of less than 1 ml/min. Following
ganisms were measured again at the end of the experiment
elution, SPEs were allowed to air-dry for 5 min. Elution of
Cipro from samples was performed stepwise by first adding
Three A. saccharum leaf disks (diameter, 1.1 cm) were
0.1 ml of 0.2 N HCl, followed by 1.5 ml of a methanol:0.2 N
added to each experimental chamber 4 d before the experiment
HCl solution and, finally, 0.4 ml of 5 mM phosphate buffer
and functioned to provide nutritional resources for L. liba.
(pH 3.0). All three elution steps were collected in the same
After 30 d, two additional leaf disks were added to each ex-
test tube. The eluent was reduced in volume to 0.8 ml, effec-
perimental chamber. Breakdown of leaf disks was not quan-
tively removing methanol from the solution and returned to a
tified in this experiment, because L. liba use leaf material for
final volume of 1.0 ml using phosphate buffer.
their cases in addition to feeding on it. Thus, loss of leaf mass
Sample analysis, based on modification of previous meth-
cannot be used as a metric for feeding activity.
ods [1,23], was performed using an Agilent 1100 series liquid
The response variable for this experiment was growth rate
chromatograph equipped with a binary pump, a Zorbax SB-
based on mass. Mass was predicted from length using a power
C18 column (4.6 ϫ 150 mm, 5-m particles), and a program-
mable fluorescence detector (Agilent, Palo Alto, CA, USA).
The elution process started with 8% acetonitrile and 92% po-
tassium phosphate buffer (5 mM, pH 3). After 2 min, the
where M ϭ mass, L ϭ length, and a and b are constants specific
percentage acetonitrile was set on a linear gradient to reach
Effects of ciprofloxacin on stream microcosms
75% at 8 min and then held steady for 3 min. Fluorescencedetection used an excitation wavelength of 278 nm and anemission wavelength of 445 nm. Five aqueous samples con-sisting of the water source used within the current study for-tified with Cipro at a concentration of either 10 or 100 g/Lwere used to validate the analytical method. Extraction effi-ciency was 107% of expected with a standard deviation of14%.
Patterns in community-level physiological profiles associ-
ated with leaf disks exposed to each Cipro concentration wereexamined with principal component analysis (PCA) using thestatistical software PC-ORD [24]. A centered variance/co-variance cross-products matrix was used, and the PCA wasnot standardized (because variables were commensurate). This
Fig. 1. Principal component analysis (PCA) of leaf disk samples within
ordination method is recommended for analysis of community-
microbial functional group space. Symbols are mean component
level physiological profile data [18] and has been used suc-
scores (n ϭ 3) for treatment groups before (closed symbols) and after
cessfully to distinguish among samples based on functional
(open symbols) 12 d of daily ciprofloxacin exposure. Carbon sources
microbial communities [25]. Before the ordination analysis,
labeled on the ends of each axis are those with the highest factorloadings for the positive or negative end of the axis. Error bars rep-
AWCD-normalized values were log (x ϩ 1)-transformed. Or-
dination axis scores were compared among treatments with ageneral-linear-model analysis of variance (ANOVA; SAS In-stitute, Cary, NC, USA). Optical density values (AWCD nor-malized) measured from carbohydrate, polymer, carboxylic
lated using a broken-stick model [24]. Specific carbon sources
acid, or amino acid carbon substrates were compared among
with the two greatest positive- and negative-factor loadings on
Cipro concentrations with a general-linear-model ANOVA.
the first two axes are shown in Figure 1.
Because four separate inferential tests were performed on the
The first axis generally was described by Cipro concentra-
same data set, a Bonferroni adjustment was used (␣ ϭ 0.013).
tion, and variation in scores was observed among leaf disk
Student-Newman-Keuls multiple-comparison tests were used
ϭ 8.9, p Ͻ 0.0002). Scores for leaf disks from
to compare means on significant ANOVAs.
chambers receiving 100 g/L of Cipro for 12 d differed from
Gammarus spp. BCI, A. saccharum leaf breakdown, and
those for all other leaf disk samples ( p Ͻ 0.05). The second
L. liba instantaneous growth rate estimates (G) were compared
axis generally was described by changes in microbial com-
among Cipro concentrations with general-linear-model AN-
munities over time, with variation in scores observed among
OVA and Student-Newman-Keuls multiple-comparison tests
ϭ 8.3, p ϭ 0.0003). On the second
(SAS Institute). Data regarding the percentage loss of leaf
axis, mean sample scores for treatment groups before Cipro
AFDM were arcsine-transformed before analyses. Bartlett’s
additions (Fig. 1, open symbols) did not differ from one an-
and Shapiro-Wilk goodness-of-fit tests indicated that BCI, G,
other ( p Ͼ 0.05), and neither did the scores for treatment
and percentage AFDM loss data had equal variances and were
groups after 12 d of Cipro additions ( p Ͼ 0.05) (Fig. 1, closed
normally distributed, thus satisfying the assumptions of AN-
symbols). Also on the second axis, mean sample scores for
the controls, 1.0, and 10.0 g/L treatments at 0 d differed frommean scores for the 12-d samples ( p Ͻ 0.05).
In terms of broader functional substrate groups, relative
microbial respiration did not differ among control and Cipro-
Leaf-associated community-level physiological profiles
treated leaf samples for carboxylic acids (F
The ordination analysis of leaf samples in functional mi-
crobial community space suggested variation in pattern among
ϭ 3.30, p ϭ 0.078) (Fig. 2). However,
Cipro concentrations after 12 d of exposure (Fig. 1), following
microbial respiration varied among Cipro treatments for car-
the interpretation that samples with similar functional micro-
ϭ 16.74, p Ͻ 0.001) (Fig. 2), with
bial communities plot near each other within the ordination
chambers receiving the 100 g/L treatment having 2.7- to 3.5-
diagram. The positioning of 0-d samples on the ordination plot
fold lower microbial respiration than controls, 1.0, and 10.0
supports this assumption, where the pattern of preconditioned
g/L treatments ( p Ͻ 0.05).
leaf disks (before receiving Cipro treatments) was similar (Fig. Detritivore growth and leaf processing
1). In addition, pattern shifts were observed relative to timeof community development (i.e., before exposure and after 12
Gammarus spp. BCI was similar among control chambers
d of exposure) for controls and all concentrations (Fig. 1). The
(1.01 Ϯ 0.04 [mean Ϯ standard error]) and those receiving
data from the 4-d exposure (not shown) followed a similar
Cipro treatments (0.1 g/L treatment, 0.97 Ϯ 0.04; 1.0 g/L
trend to that from the 12-d exposure but had less separation
ϭ 1.25, p ϭ 0.300) (Fig. 3).
of points in the ordination diagram. For the ordination con-
Percentage loss of A. saccharum AFDM was similar among
taining the 0- and 12-d data, the cumulative variance explained
the control (62.1% Ϯ 1.21%) and Cipro treatments (0.1 g/L
by the first four axes of the PCA, was 72.4%, with 32.4 and
treatment, 59.3% Ϯ 2.65%; 1.0 g/L treatment, 61.5% Ϯ
15.3% attributed to the first and second axes, respectively.
ϭ 0.44, p ϭ 0.650) (Fig. 3). Lepidostoma liba
Axes 5 and higher had eigenvalues greater than those calcu-
instantaneous growth rate estimates (G) were similar among
Fig. 3. Mean (Ϯ standard error) (A) Gammarus spp. body condition index (BCI) and (B) percentage loss of Acer saccharum ash-free dry mass (AFDM) after 30 d of ciprofloxacin exposure. For the BCI, a value of one indicates the expected organism AFDM based on length from the linear-regression model. Sample sizes (i.e., experimental chambers) for control, 0.1, and 1.0 g/L treatments were 12, 9, and 11, respectively.
controls (0.0094 Ϯ 0.0016) and chambers receiving Ciprotreatments (10 g/L treatment, 0.0091 Ϯ 0.0011; 100 g/Ltreatment, 0.0095 Ϯ 0.0017; F
ϭ 0.01, p ϭ 0.987) (Fig. 4).
Survival across treatments for L. liba was 90.4, 91.7, and
89.4% based on the total recovered for control, 10.0, and 100.0g/L treatments, respectively. Because L. liba growth esti-mates were based on averages of three individuals per exper-imental unit (i.e., individuals were not tracked), growth anal-yses were only conducted on chambers in which all three in-dividuals were recovered alive. As a result, analyses wereconducted on 12, 10, and 9 replicate chambers for the control,10, and 100 g/L treatments, respectively. Survival acrosstreatments for Gammarus spp. was 86.7, 95.6, and 94.5% forcontrol, 0.1, and 1.0 g/L treatments, respectively. The subsetof Gammarus spp. collected at the start of the experiment hada higher BCI (1.25 Ϯ 0.05; i.e., based on the regression equa-tion of experimental control organisms only) than did the con-trols and both Cipro treatments after 30 d, which may havebeen caused by the onset of reproduction during the experi-
Fig. 2. Mean (Ϯ standard error) microbial respiration (i.e., optical density
normalized for average well color development) for EcoPlate carbonsubstrates that were polymers, carbohydrates, amino acids, or carboxylic
Water chemistry and Cipro stability
acids. Within each carbon substrate group, ciprofloxacin concentrationsthat share a common letter above their error bar are similar ( p Ͼ 0.05).
Water chemistry in the Gammarus spp. experiment varied
Sample sizes (i.e., experimental chambers) for the control and each con-
little because of storage in the laboratory and within experi-
Effects of ciprofloxacin on stream microcosms
and clustered together. After a period of 12 d, all leaf-asso-ciated communities shifted on the second axis, likely attrib-utable to successional processes of the community such as thatobserved by Bundy et al. [26]. This idea is further supportedby the shift in pattern and relative positions of the control and1.0 g/L samples at both time points (0 and 12 d). After theCipro exposure period, variation in the function of commu-nities on leaf samples occurred, primarily along the first or-dination axis. These results indicate that repeated microcosmexposures of 100 g/L altered leaf-associated microbial com-munities. Samples from the 10 g/L treatment also were spa-tially shifted along the first ordination axis. Although the spa-tial patterning may suggest a dose–response pattern, the 10g/L treatment was not statistically different from the 1 g/L treatment or the controls. Community-level physiologicalprofiles have been used previously to evaluate impacts of con-
Fig. 4. Mean (Ϯ standard error) Lepidostoma liba daily instantaneous
taminants, such as diesel fuel and methyl bromide, in the en-
growth rate after 45 d of ciprofloxacin exposure. Sample sizes (i.e.,
vironment [26,27]. However, to our knowledge, this is the first
experimental chambers) for control, 10.0, and 100.0 g/L treatmentswere 12, 10, and 9, respectively.
report using this technique to evaluate potential impacts ofantibiotics to stream microbial communities.
Much of the variation in carbon substrate utilization among
mental chambers 24 h postrenewal. Ranges for parameters
leaf samples was explained by the respiration of carbohydrates.
measured in chambers over the entire 30-d experiment were
This was the only substrate type on the EcoPlates that varied
as follows: temperature, 13.0–8.9ЊC; dissolved oxygen, 13.3–
among Cipro treatments. Other studies have shown differential
10.5 mg/L; pH 8.1–7.0; and conductivity, 455–273 S/cm.
respiration among EcoPlate carbon substrates by microbial
The variation in conductivity was attributed to higher flow
communities [28], and the variation was attributed to the type
conditions during one collection period because of snow melt
of sample. In the current study, similar samples shifted res-
and rain events. Over the 45-d L. liba experiment, water-chem-
piration patterns within a particular group of substrates (i.e.,
istry parameters were relatively consistent with the following
carbohydrates) because of the addition of Cipro, with chambers
ranges: temperature, 14.9–13.6ЊC; dissolved oxygen, 10.7–8.4
receiving 100 g/L having a reduced measure of respiration.
mg/L; pH 8.4–8.1; and conductivity, 428–389 S/cm. The
Primarily driving this pattern were three carbohydrates: -
addition of acetic acid and Cipro to experimental chambers
methyl-D-glucoside, D-mannitol, and N-acetyl-D-glucosamine.
did not have an effect on pH (control: mean, 8.29; 100 g/L
The ordination results support this finding with -methyl-D-
treatment: mean, 8.33; t ϭ Ϫ1.13, p ϭ 0.269). Final water
glucoside having the greatest positive loading on the first axis,
chemistry was not measured in the microbial community ex-
indicating that respiration of this substrate was more closely
periments; however, water renewals for this experiment con-
associated with the control, 1.0, and 10.0 g/L samples.
sisted of the same source of control and Cipro-treated water
The variation in optical density (i.e., a measure of microbial
as that used in the L. liba experiment (i.e., they were run
respiration) for carbohydrate substrates suggested that average
concurrently). Water chemistry of prepared solutions was sim-
microbial respiration among these substrates was reduced for
ilar to that measured within the experimental chambers after
leaf disk samples exposed to 100 g/L for 12 d. This indicates
a reduction in some carbohydrate-using microbes that may be
Cipro concentrations within the test systems declined com-
useful to detritivores for metabolizing carbohydrates or pro-
pared to the renewal solution; however, the solution concen-
viding exogenous enzymes. Despite this, growth and condition
trations within the test chambers were relatively stable. The
index were not variable among treatments for both detritivores
average concentration and standard deviation 1 h after renewal
examined, even for L. liba exposed to 100 g/L of Cipro. One
was 83 Ϯ 9 g/L for the 100 g/L treatments and 6.6 Ϯ 1.5
possible explanation is that some detritivores may produce
g/L for the 10 g/L treatments. After 24 h, just previous to
enzymes endogenously and digest carbohydrates, such as cel-
the next renewal, the concentrations were 63 Ϯ 20 and 5.1 Ϯ
lulose [7]. This has been reported in Gammarus pulex, in
1.4 g/L for the 100 and 10 g/L treatments, respectively.
which digestive fluids from the gut contained cellulases that
Thus, average fluctuations in concentration were 25% or less
hydrolyzed multiple cellulose substrates [29], and in Gam-
between renewals based on changes from 1 to 24 h postre-
marus lacustris, in which endoglucanase and -glucosidase
newal. Mean exposure concentration, assuming a linear decline
were found to occur endogenously [30].
in Cipro concentration, was 73 Ϯ 14 and 5.9 Ϯ 1.2 g/L forthe 100 and 10 g/L treatments, respectively. Detritivore growth indices and leaf processingDISCUSSION
Detritivores are a critical functional component in many
stream ecosystems because of their roles in the processing of
Leaf-associated community level physiological profiles
allochthonous energy sources and nutrient cycling. Microbial
Results of the ordination analysis indicate variation in the
communities are important in these processes, because they
functional microbial community both through time and among
can act directly as a food resource associated with detritus [31]
Cipro concentrations, similar to the separation observed among
and as endosymbionts providing enzymes for metabolism of
aqueous microbial communities from sites of varying envi-
cellulose and refractory leaf constituents. It is believed such
ronmental conditions [17]. The initial communities (i.e., before
enzymes do not occur endogenously in many aquatic inver-
Cipro exposures) were similar among experimental chambers
tebrates [12]. Considering that microbes are critical for ac-
quisition of nutrition for detritivores, it was predicted that a
able to capitalize on this redundancy or rely more heavily on
microcosm exposure to an antibiotic would affect microbes
fungi that are not within the spectrum of activity of Cipro.
and, in turn, influence responses indicative of reduced resource
Finally, the species examined here may endogenously pro-
acquisition, such as detritivore growth and body condition.
duce enzymes that enable them to digest detritus and leaf
However, results from the present study indicate that L. liba
material and to rely little on endosymbionts. Evidence suggests
growth rate and condition of Gammarus spp. were not influ-
that this may occur in some Gammarids [29,30,37]. Alterna-
enced by the Cipro exposure concentrations tested. Lepido-
tively, in other leaf-shredding stream invertebrates, microbial
stoma liba growth was observed in all treatment groups, and
enzymes play an important role, either expressed from en-
the rate of growth did not vary among controls and Cipro-
dosymbionts in the gut (e.g., Tipula abdominalis) or via in-
treated microcosms. Growth rates observed in these experi-
gestion of microbial-conditioned leaf material (e.g., Pteron-
ments were slightly higher than those reported for L. quercinaarcys proteus) [12]. Both the degree that L. liba depend on
fed red alder (Alnus rubra) leaves at 15ЊC.
microbial-derived enzymes and the mechanism of obtaining
Growth responses of stream-dwelling caddisflies have long
those enzymes (i.e., endosymbiont or via conditioned leaves)
been used as a sensitive indicator of stream resource condition
in laboratory bioassays. For example, growth and condition
Detritivore leaf processing also provides information re-
of Olinga feredayi and growth rates of Psychoglypha spp.
garding environmental factors in an ecosystem context and is
were related to diets of varying bacterial, fungal, and algal
related to pH gradients [38], detritivore density [39], and the
densities [8,32], growth rates of L. quercina were limited by
type and composition of leaves present in streams [40]. De-
a lack of high-quality food [33], and differential growth of
tritivores preferentially feed on leaves conditioned by bacteria
Pycnopsyche spp. was observed among treatments of different
and fungi, because it is believed these organisms make leaf
leaf species [34]. Thus, variation in L. liba growth rates would
material more palatable via their enzymes [12,41]. Increasedpalatability facilitates feeding and should result in increased
be likely if an impact of Cipro additions on nutritional quality
processing rates [8,41]. The similarity of loss of leaf AFDM
of leaf disks occurred in the current study.
among Cipro treatments in the current study suggests that
Several explanations are possible for the lack of effect of
Gammarus spp. did not alter leaf processing in response to
Cipro on detritivore growth and condition endpoints. First,
potentially reduced palatability. Furthermore, it also may be
Cipro concentrations may have been below the levels that
hypothesized that Gammarus processing of Cipro-treated
substantially reduce the symbiotic function and biomass of
leaves would increase in response to a reduction in nutritional
microbes. To our knowledge, a dose–response relationship be-
quality per unit area of leaf disk (i.e., reduced microbial pop-
tween biomass or functioning of gut bacteria and any antibiotic
ulations). A pattern suggesting this alternative also was not
has not been reported, but such a response may require high
observed. These results suggest that Cipro treatment did not
antibiotic concentrations. For example, in a previous study in
influence microbial activity in a way that could be detected
which gut microbes were to be eliminated in Gammarus la-
by detritivore processing of leaf material. custris, McGrath and Matthews [30] used an antibiotic mixtureof tetracycline, streptomycin, nystatin, and metronidazole at
Environmental concentrations and effects of Cipro
concentrations high enough (mg/L level) to treat diseased fishin an aquaculture setting. In terms of effective Cipro concen-
The nominal and measured exposure concentrations used
in these experiments were above those typically reported for
trations, another unknown is the concentration necessary to
streams and rivers. Mean aqueous concentrations measured in
penetrate the hepatopancreas and other glands and structures
river water (Glatt River, Switzerland) ranged from 0.008 to
of the digestive tract, where endosymbionts typically reside.
0.015 g/L [1], and the median concentration detected in U.S.
In the Gammarus experiment, the initial exposure concentra-
streams was 0.020 g/L [2]. Although the 0.1 and 1.0 g/L
tions were high enough to substantially reduce some bacterial
concentrations of Cipro used in the current study were less
populations [6]. However, in our experimental chambers, the
than the effective concentration for growth in the cyanobacteria
complex matrix of water, sediment, and leaf disks may have
Microcystis aeruginosa (EC50 ϭ 17 g/L [3]), a Cipro con-
included factors such as dissolved and particulate organic mat-
centration of 0.5 g/L reduced Pseudomonas aeruginosa and
ter that reduced availability of Cipro. Future work should ex-
Escherichia coli culture viability to 0.01% of controls, and an
amine the role of these factors in the toxicity of antibiotics to
exposure concentration of 0.1 g/L resulted in a 10-fold re-
better understand potential impacts to stream ecosystem end-
duction in culture viability [6]. Thus, selected exposure con-
centrations had the potential to impact stream microbial pro-
Second, there may be sufficient functional redundancy
cesses, and the 0.1 g/L treatment may have approached con-
within microbial communities in this system such that mi-
centrations observed in aquatic systems. However, dramatic
crobes (i.e., fungi) not within the spectrum of activity for Cipro
shifts in community-level physiological profiles of microbes
persisted, providing a continued source of nutrition and/or ex-
were only observed in the 100 g/L treatment. Although the
ogenous enzymes for detritivores. This may be the case par-
acute toxicity of Cipro to Gammarus spp. and L. liba is un-
ticularly for the L. liba experiment involving a 100 g/L treat-
known, the concentrations used in these experiments likely
ment. Several researchers have explored functional redundan-
were well below levels that would elicit acute effects [3];
cy in microbial communities [35,36]. In particular, Wohl et al.
reported Cipro no-observed-effect concentrations for the zoo-
[35] showed redundancy in cellulose degraders in a riverine
plankton Daphnia magna were greater than 10 mg/L [3,42].
ecosystem. The observations that 100 g/L treatments did,
Few studies regarding antibiotics have examined indirect
indeed, influence bacterial communities (as suggested by the
ecological effects among species and trophic levels (i.e., com-
EcoPlate data) and that L. liba growth was unaffected lend
munity- and ecosystem-level responses). Richards et al. [43]
support to the idea of redundancy in stream microbial com-
examined the response of zooplankton community structure
munities [35] and provide possible evidence of detritivores
and function to a combination of pharmaceuticals that included
Effects of ciprofloxacin on stream microcosms
Cipro. With increasing Cipro concentration, diversity of zoo-
the impact of Cipro alone to stream detritivores and related
plankton taxa was reduced, and abundance was elevated. How-
microbial communities at environmentally relevant concentra-
ever, only the mixture was examined, so effects caused by
tions is probably low. However, only two detritivores and a
Cipro alone were unknown. In another study, zooplankton bio-
limited component of the microbial community were exam-
mass in microcosms was similar among controls and those
ined. It is possible that some stream detritivores or other aquat-
treated with a mixture of four tetracyclines [44]. The results
ic species having specialized relationships with stream mi-
of the current study concur with the biomass finding. However,
crobes might be more sensitive. Future studies should examine
it should also be noted that in the current study, stream de-
other taxa and microbially mediated ecosystem processes, such
tritivores were examined (i.e., species that may have a very
as denitrification and nitrification, for identifying the risk of
different relationship with microbes than zooplankton species
Acknowledgement—This research was supported by U.S. Environ-
mental Protection Agency Science To Achieve Results (STAR) grant
Sources of variation in the current study included factors
R82900801-0. We thank F. Wilhelm, A. Trimble, and D. Walther for
that may have influenced measures of mass for Gammarus
spp., as suggested by a reduction in BCI when compared to
REFERENCES
initial organisms. The reproductive period for Gammarus spp.
1. Golet EM, Alder AC, Giger W. 2002. Environmental exposure
in Big Creek coincided with the timing of this experiment (D.
and risk assessment of fluoroquinolone antibacterial agents in
Walther and M.R. Whiles, Southern Illinois University, Car-
wastewater and river water of the Glatt Valley Watershed, Swit-
bondale, IL, USA, unpublished data), and indeed, reproduction
zerland. Environ Sci Technol 36:3645–3651.
2. Kolpin DW, Furlong ET, Meyer MT, Thurman EM, Zaugg SD,
of Gammarus spp. was observed in many of the experimental
Barber LB, Buxton HT. 2002. Pharmaceuticals, hormones, and
chambers after 20 d and may have contributed to the overall
other organic wastewater contaminants in U.S. streams, 1999–
reduction in BCI across treatments. Despite this, it was im-
2000: A national reconnaissance. Environ Sci Technol 36:1202–
portant to examine the organisms and stream matrices asso-
ciated with this part of the annual cycle, because it is a period
3. Robinson AA, Belden JB, Lydy MJ. 2005. Toxicity of fluoro-
quinolone antibiotics to aquatic organisms. Environ Toxicol Chem
when Cipro levels typically are more elevated [1].
In addition, examining community-level physiological pro-
4. Suberkropp K, Klug MJ. 1976. Fungi and bacteria associated with
files using carbon substrates only provides information re-
leaves during processing in a woodland stream. Ecology 57:707–
garding a component of the overall microbial community. Mi-
5. Wolfson JS, Hooper DC. 1985. The fluoroquinolones: Structures,
crobes, such as fermenting and anaerobic bacteria, as well as
mechanisms of action and resistance, and spectra of activity in
fungi are very important to the stream ecosystem, but they
vitro. Antimicrob Agents Chemother 28:581–586.
may not be detected using this carbon source utilization meth-
6. Chalkley LJ, Koornhof HJ. 1985. Antimicrobial activity of cip-
od. Additional techniques, such as using whole-community
rofloxacin against Pseudomonas aeruginosa, Escherichia coli,
DNA extracts or measuring phospholipid fatty acids, may pro-
and Staphylococcus aureus determined by the killing curve meth-od: Antibiotic comparisons and synergistic interactions. Antimi-
vide additional insight regarding impacts of antibiotics on
crob Agents Chemother 28:331–342.
stream microbial communities [18]. Despite this, sufficient in-
7. Harris JM. 1993. The presence, nature, and role of gut microflora
formation to successfully discriminate among microbial com-
in aquatic invertebrates: A synthesis. Microb Ecol 25:195–231.
munities may be obtained using the carbon substrate technique
8. Arsuffi TL, Suberkropp K. 1986. Growth of two stream caddisflies
(Trichoptera) on leaves colonized by different fungal species. J
[18]. In addition, several comparative studies have indicated
North Am Benthol Soc 5:297–305.
similar results between carbon substrate utilization and other
9. Webster JR, Benfield EF, Ehrman TP, Schaeffer MA, Tank JL,
Hutchens JJ, D’Angelo DJ. 1999. What happens to allochthonous
Measured Cipro concentrations in the current study indi-
material that falls into streams? A synthesis of new and published
cated that Cipro ranged from 51 to 83% of nominal concen-
information from Coweeta. Freshw Biol 41:687–705.
10. Cummins KW. 1973. Trophic relations of aquatic insects. Annu
trations. Although the exposures may have been reduced by a
factor of two, the exposure concentrations were still well above
11. Hall RO, Meyer JL. 1998. The trophic significance of bacteria in
environmentally relevant concentrations. Furthermore, the loss
a detritus-based stream food web. Ecology 79:1995–2012.
of Cipro was likely caused by sorption to organic matter within
12. Sinsabaugh RL, Linkins AE, Benfield EF. 1985. Cellulose di-
gestion and assimilation by three leaf-shredding aquatic insects.
the test system. Thus, although a Cipro concentration on leaf
material was not measured, the leaf material may have had
13. Wallace JB, Eggert SL, Meyer JL, Webster JR. 1997. Multiple
much higher concentrations than test water. Future research
trophic levels of a forest stream linked to terrestrial litter inputs.
should evaluate the implications of Cipro partitioning to or-
14. MacNeil C, Dick JTA, Elwood RW. 1997. The trophic ecology
of freshwater Gammarus spp. (Crustacea: Amphipoda): Problemsand perspectives concerning the functional feeding group concept. CONCLUSIONS
Results of the current study suggest that repeated micro-
15. Minshall GW, Petersen RC, Bott TL, Cushing CE, Cummins KW,
cosm exposures to Cipro at 100 g/L and, to a lesser degree,
Vannote RL, Sedell JR. 1992. Stream ecosystem dynamics of theSalmon River, Idaho—An eighth-order system. J North Am Ben-
at 10 g/L altered the function of leaf-associated microbial
communities. However, detritivore growth indices and leaf
16. Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing
breakdown were not affected at these and lower concentrations.
CE. 1980. The river continuum concept. Can J Fish Aquat Sci
Furthermore, Cipro exposures at concentrations at or less than
17. Garland JL, Mills AL. 1991. Classification and characterization
1.0 g/L did not appear to influence leaf microbial commu-
of heterotrophic microbial communities on the basis of patterns
nities. Concentrations used in these experiments were above
of community level sole-carbon-source utilization. Appl Environ
those reported in stream systems; thus, our results suggest that
18. Mills AL, Garland JL. 2002. Application of physiological profiles
sponses to algal and bacterial manipulations in streams. Hydro-
to assessment of community properties. In Hurst CJ, Crawford
RL, Knudson GR, McInerney MJ, Stetzenbach LD, eds, Manual
32. Burrell GP, Ledger ME. 2003. Growth of a stream-dwelling cad-
of Environmental Microbiology. ASM, Washington, DC, pp 135–
disfly (Olinga feredayi: Conoesucidae) on surface and hyporrheic
food resources. J North Am Benthol Soc 22:92–104.
19. Green AJ. 2001. Mass/length residuals: Measures of body con-
33. Grafius E, Anderson NH. 1979. Population dynamics, bioener-
dition or generators of spurious results. Ecology 82:1473–1483.
getics, and role of Lepidostoma quercina Ross (Trichoptera: Lep-
20. Legendre P, Legendre L. 1998. Numerical Ecology. Elsevier, Am-
idostomatidae) in an Oregon woodland stream. Ecology 60:433–
21. Benke AC, Huryn AD, Smock LA, Wallace JB. 1999. Length–
34. Hutchens JJ, Jr., Benfield EF, Webster JR. 1997. Diet and growth
of a leaf-shredding caddisfly in southern Appalachian streams of
mass relationships for freshwater macroinvertebrates in North
contrasting disturbance history. Hydrobiologia 346:193–201.
America with particular reference to the southeastern United
35. Wohl DL, Arora S, Gladstone JR. 2004. Functional redundancy
States. J North Am Benthol Soc 18:308–343.
supports biodiversity and ecosystem function in a closed and
22. Benke AC. 1996. Secondary production of macroinvertebrates.
constant environment. Ecology 85:1534–1540.
In Hauer FR, Lamberti GA, eds, Methods in Stream Ecology.
36. Mikola J, Seta¨la¨ H. 1998. Relating species diversity to ecosystem
Academic, San Diego, CA, USA, pp 557–578.
functioning: Mechanistic backgrounds and experimental approach
23. Yorke JC, Froc P. 2000. Quantitation of nine quinolones in chick-
with a decomposer food web. Oikos 83:180–194.
en tissues by high-performance liquid chromatography with fluo-
37. Zimmer M, Bartholme S. 2003. Bacterial endosymbionts in Asel-
rescence detection. J Chromatogr A 882:63–77. lus aquaticus (Isopoda) and Gammarus pulex (Amphipoda) and
24. McCune B, Mefford MJ. 1999. PC-ORD. Multivariate Analysis
their contribution to digestion. Limnol Oceanogr 48:2208–2213. of Ecological Data, Version 4. MjM Software Design, Gleneden
38. Dangles O, Gessner MO, Guerold F, Chauvet E. 2004. Impacts
of stream acidification on litter breakdown: Implications for as-
25. Garland JL. 1996. Analytical approaches to the characterization
sessing ecosystem functioning. J Anim Ecol 41:365–378.
of samples of microbial communities using patterns of potential
39. Fazi S, Rossi L. 2000. Effects of macrodetritivores density on
C source utilization. Soil Biol Biochem 28:213–221.
leaf detritus processing rate: A macrocosm experiment. Hydro-
26. Bundy JG, Paton GI, Campbell CD. 2002. Microbial communities
in different soil types do not converge after diesel contamination.
40. Swan CM, Palmer MA. 2004. Leaf diversity alters litter break-
J Appl Microbiol 92:276–288.
down in a Piedmont stream. J North Am Benthol Soc 23:15–28.
27. Ibekwe AM, Papiernik SK, Gan J, Yates SR, Yang C-H, Crowley
41. Graca MAS. 2001. The role of invertebrates on leaf litter decom-
DE. 2001. Impact of fumigants on soil microbial communities.
position in streams—A review. Int Rev Hydrobiol 86:383–393. Appl Environ Microbiol 67:3245–3257.
42. Halling-Sorensen B, Holten Lutzhoft H-C, Andersen HR, Inger-
slev F. 2000. Environmental risk assessment of antibiotics: Com-
28. Lehman RM, Colwell FS, Ringelberg DB, White DC. 1995. Com-
parison of mecillinam, trimethoprim, and ciprofloxacin. J Anti-
bined microbial community-level analysis for quality assurance
of terrestrial subsurface cores. J Microbiol Methods 22:263–281.
43. Richards SM, Wilson CJ, Johnson DJ, Castle DM, Lam M, Ma-
29. Monk DC. 1977. The digestion of cellulose and other dietary
bury SA, Sibley PK, Solomon KR. 2004. Effects of pharmaceu-
components, and pH of the gut in the amphipod Gammarus pulex
tical mixtures in aquatic microcosms. Environ Toxicol Chem 23:
(L.). Freshw Biol 7:31–440.
30. McGrath CC, Matthews RA. 2000. Cellulase activity in the fresh-
44. Wilson CJ, Brain RA, Sanderson H, Johnson DJ, Bestari KT,
water amphipod Gammarus lacustris. J North Am Benthol Soc
Sibley PK, Solomon KR. 2004. Structural and functional re-
sponses of plankton to a mixture of four tetracyclines in aquatic
31. Fuller RL, Kennedy BP, Nielson C. 2004. Macroinvertebrate re-
microcosms. Environ Sci Technol 38:6430–6439.
VIDEX - General User Guidelines Dear User, Welcome to VIDEX, the software of the Federal Republic of Germany which enables you to complete your application form for a Schengen visa online. We want to make VIDEX as easy as possible, so here’s all the relevant information on how to use VIDEX and how to submit your application. Please read the following information carefully bef
Avis juridique important 31976L0768 Richtlinie 76/768/EWG des Rates vom 27. Juli 1976 zur Angleichung der Rechtsvorschriften der Mitgliedstaaten über kosmetische Mittel Amtsblatt Nr. L 262 vom 27/09/1976 S. 0169 - 0200 Griechische Sonderausgabe: Kapitel 13 Band 4 S. 0145 Spanische Sonderausgabe: Kapitel 15 Band 1 S. 0206 Portugiesische Sonderausgabe: Kapitel 15 Band 1 S. 0206 Fi
 Environmental Toxicology and Chemistry, Vol. 25, No. 6, pp. 1598–1606, 2006
EFFECTS OF THE ANTIBIOTIC CIPROFLOXACIN ON STREAM MICROBIAL
COMMUNITIES AND DETRITIVOROUS MACROINVERTEBRATES
JONATHAN D. MAUL,† LANCE J. SCHULER,† JASON B. BELDEN,† MATT R. WHILES,‡ and MICHAEL J. LYDY*†
†Fisheries and Illinois Aquaculture Center and Department of Zoology, ‡Department of Zoology,
Southern Illinois University, Carbondale, Illinois 62901, USA
(Received 25 July 2005; Accepted 10 November 2005)
Abstract—Microbial communities play an important role in stream ecosystem processes, such as breakdown of senescent leaf litter,
Environmental Toxicology and Chemistry, Vol. 25, No. 6, pp. 1598–1606, 2006
EFFECTS OF THE ANTIBIOTIC CIPROFLOXACIN ON STREAM MICROBIAL
COMMUNITIES AND DETRITIVOROUS MACROINVERTEBRATES
JONATHAN D. MAUL,† LANCE J. SCHULER,† JASON B. BELDEN,† MATT R. WHILES,‡ and MICHAEL J. LYDY*†
†Fisheries and Illinois Aquaculture Center and Department of Zoology, ‡Department of Zoology,
Southern Illinois University, Carbondale, Illinois 62901, USA
(Received 25 July 2005; Accepted 10 November 2005)
Abstract—Microbial communities play an important role in stream ecosystem processes, such as breakdown of senescent leaf litter,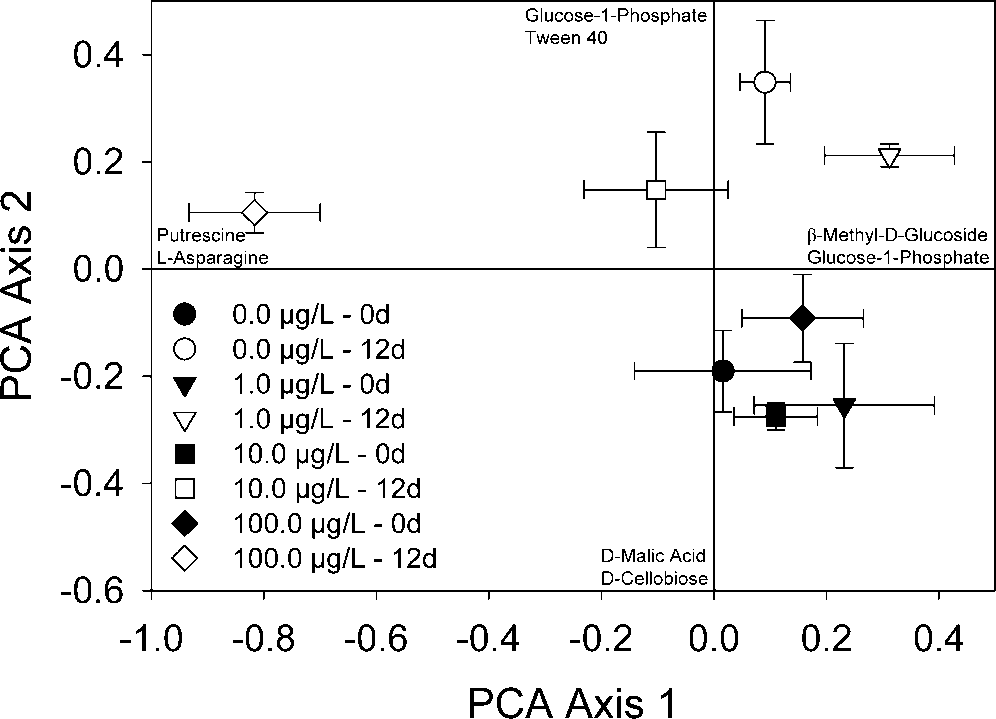 Effects of ciprofloxacin on stream microcosms
75% at 8 min and then held steady for 3 min. Fluorescencedetection used an excitation wavelength of 278 nm and anemission wavelength of 445 nm. Five aqueous samples con-sisting of the water source used within the current study for-tified with Cipro at a concentration of either 10 or 100 g/Lwere used to validate the analytical method. Extraction effi-ciency was 107% of expected with a standard deviation of14%.
Effects of ciprofloxacin on stream microcosms
75% at 8 min and then held steady for 3 min. Fluorescencedetection used an excitation wavelength of 278 nm and anemission wavelength of 445 nm. Five aqueous samples con-sisting of the water source used within the current study for-tified with Cipro at a concentration of either 10 or 100 g/Lwere used to validate the analytical method. Extraction effi-ciency was 107% of expected with a standard deviation of14%.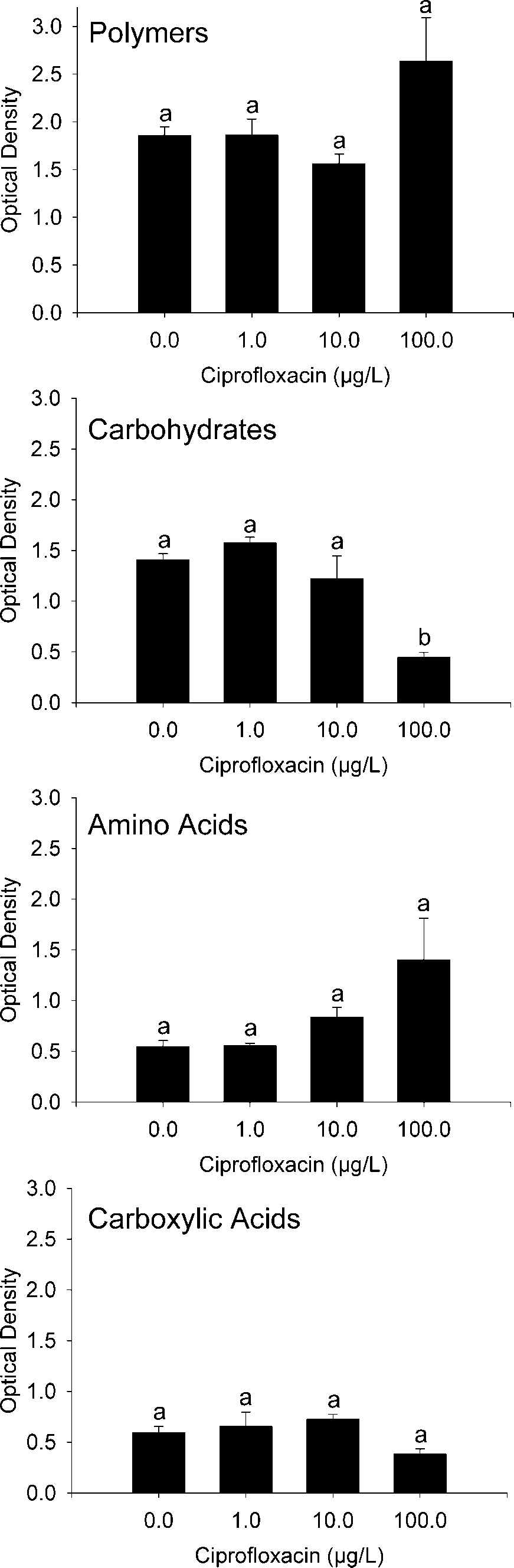
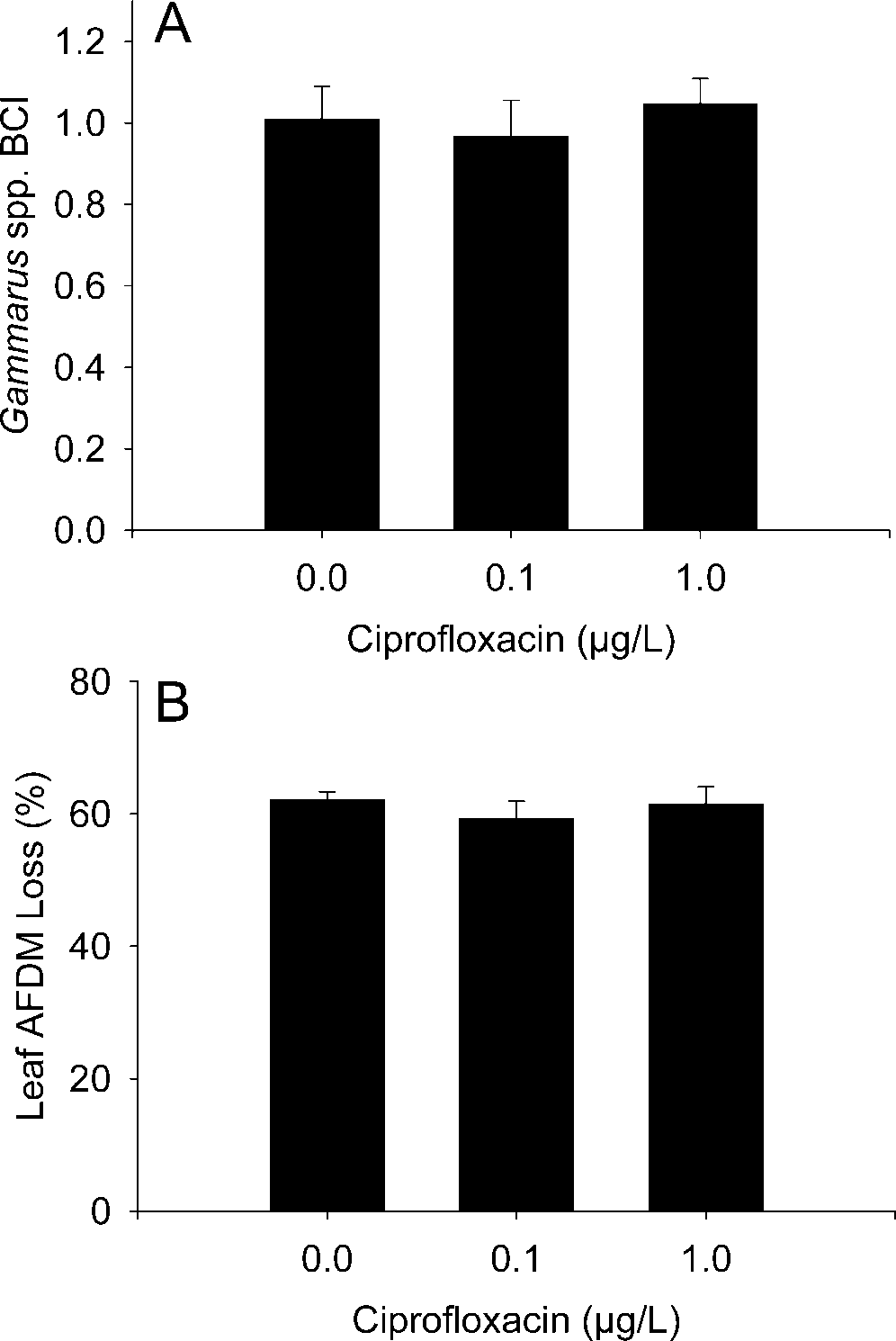 Fig. 3. Mean (Ϯ standard error) (A) Gammarus spp. body condition
Fig. 3. Mean (Ϯ standard error) (A) Gammarus spp. body condition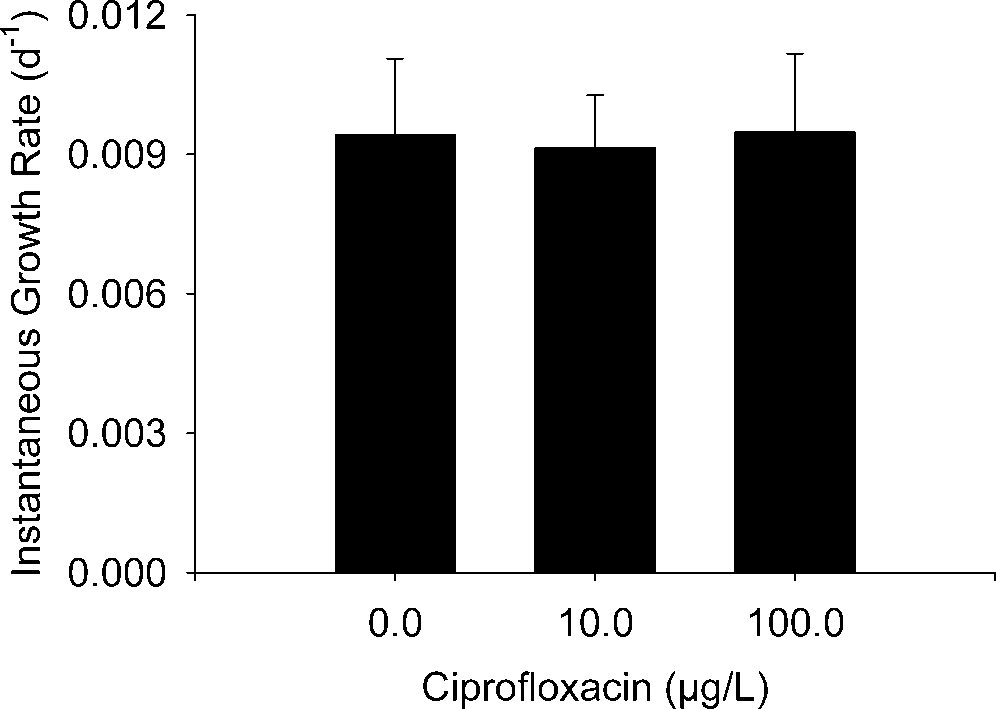 Effects of ciprofloxacin on stream microcosms
and clustered together. After a period of 12 d, all leaf-asso-ciated communities shifted on the second axis, likely attrib-utable to successional processes of the community such as thatobserved by Bundy et al. [26]. This idea is further supportedby the shift in pattern and relative positions of the control and1.0 g/L samples at both time points (0 and 12 d). After theCipro exposure period, variation in the function of commu-nities on leaf samples occurred, primarily along the first or-dination axis. These results indicate that repeated microcosmexposures of 100 g/L altered leaf-associated microbial com-munities. Samples from the 10 g/L treatment also were spa-tially shifted along the first ordination axis. Although the spa-tial patterning may suggest a dose–response pattern, the 10g/L treatment was not statistically different from the 1 g/L treatment or the controls. Community-level physiologicalprofiles have been used previously to evaluate impacts of con-
Fig. 4. Mean (Ϯ standard error) Lepidostoma liba daily instantaneous
taminants, such as diesel fuel and methyl bromide, in the en-
growth rate after 45 d of ciprofloxacin exposure. Sample sizes (i.e.,
vironment [26,27]. However, to our knowledge, this is the first
experimental chambers) for control, 10.0, and 100.0 g/L treatmentswere 12, 10, and 9, respectively.
Effects of ciprofloxacin on stream microcosms
and clustered together. After a period of 12 d, all leaf-asso-ciated communities shifted on the second axis, likely attrib-utable to successional processes of the community such as thatobserved by Bundy et al. [26]. This idea is further supportedby the shift in pattern and relative positions of the control and1.0 g/L samples at both time points (0 and 12 d). After theCipro exposure period, variation in the function of commu-nities on leaf samples occurred, primarily along the first or-dination axis. These results indicate that repeated microcosmexposures of 100 g/L altered leaf-associated microbial com-munities. Samples from the 10 g/L treatment also were spa-tially shifted along the first ordination axis. Although the spa-tial patterning may suggest a dose–response pattern, the 10g/L treatment was not statistically different from the 1 g/L treatment or the controls. Community-level physiologicalprofiles have been used previously to evaluate impacts of con-
Fig. 4. Mean (Ϯ standard error) Lepidostoma liba daily instantaneous
taminants, such as diesel fuel and methyl bromide, in the en-
growth rate after 45 d of ciprofloxacin exposure. Sample sizes (i.e.,
vironment [26,27]. However, to our knowledge, this is the first
experimental chambers) for control, 10.0, and 100.0 g/L treatmentswere 12, 10, and 9, respectively.