Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Mcconnaughey, ted a., walter h. adey, and allegra m. small community and environmental influences on reef coral calcification
Limnol. Oceanogr., 45(7), 2000, 1667–1671
᭧ 2000, by the American Society of Limnology and Oceanography, Inc.
Community and environmental influences on reef coral calcification
Abstract—Reef corals calcify faster in the presence of non-
1E), the calcifier should fix carbon almost twice as efficient-
calcareous algae. Ratios of calcification to photosynthesis ap-
ly. Figure 2B and C explore how photosynthesis, calcifica-
pear to be affected by the ratio of alkalinity to acidity, which
tion, and diffusion interact to control chemistry at the au-
controls how efficiently the protons from calcification convert
totroph’s absorptive surface. With reduced diffusion
bicarbonate to carbon dioxide. By forcing calcifiers to calcify
problems, calcareous plants and algae–invertebrate symbio-
faster, algal proliferation on nutrient-enriched reefs may ad-
ses often adopt massive morphologies.
Gattuso et al. (2000) argued that calcification does not
help photosynthesis, based on observations that a coral in-cubated at reduced Ca2ϩ concentrations (2.85 mMol kgϪ1)
Reef corals and calcareous algae usually calcify fastest
maintained normal photosynthesis but calcified less. Calcium
during the daytime and often fix carbon into calcium car-
transport inhibitors and Ca2ϩ concentrations below 0.5 mM
bonate and biomass at similar rates. Light-enhanced calcifi-
have, however, inhibited photosynthesis in other experiments
cation has long been recognized (Kawaguti and Sakumoto
(reviewed by McConnaughey and Whelan 1997). The dis-
1949; Goreau 1959) but still poses a chicken-and-egg dilem-
parity may depend on how strongly the treatment inhibits
ma: Does photosynthesis stimulate calcification by raising
the proton flux from the calcifying space. This Hϩ flux ap-
CO ϭ concentrations and the CaCO saturation state of the
pears to be coupled to Ca2ϩ fluxes through Ca2ϩ/2Hϩ ex-
change catalyzed by the enzyme Ca2ϩ adenosine triphospha-
thereby stimulate photosynthesis? Does photosynthesis pro-
tase (ATPase), as illustrated in Fig. 1C. Gattuso’s Ca2ϩ
vide metabolic energy to power calcification? We examine
concentrations were adequate to support this coupling;
these questions and their extension from the organismal to
hence, Hϩ fluxes probably continued, supported temporarily
by OHϪ and CO ϭ accumulation within the calcifying space.
CO diffusion through the boundary layer surrounding an
Gattuso’s experiments therefore demonstrate that photosyn-
aquatic autotroph (Fig. 1A) can support photosynthetic rates
thesis does not depend on CaCO precipitation, but they do
of about 0.2 mol mϪ2 sϪ1, calculated using Fick’s first law,
not rule out stimulation by the Hϩ (and Ca2ϩ ) fluxes that
with a dissolved molecular CO concentration of 10 Mol
produce calcification under normal conditions. In light of
kgϪ1, a 50-m–thick boundary layer, and 50% depletion of
typical Michaelis curves for photosynthesis (Fig. 1E) and
CO level at the autotroph’s absorptive surface. Algae and
calcification’s ability to counteract CO depletion (Fig. 2), it
corals often have photosynthetic rates that are several times
would be surprising if calcification did not sometimes benefit
that fast, indicating that they use mainly bicarbonate. How-
ever, bicarbonate utilization requires additional protons (Hϩ
Calcification and photosynthesis become coupled through
ϩ HCO Ϫ ϭ CH O ϩ O ). The protons may derive from
H O and HCO Ϫ, with a corresponding efflux of OHϪ and
CO ϭ (Fig. 1B). Large OHϪ and CO ϭ effluxes imply alka-
linization and therefore CO depletion at the absorptive sur-
face. This process reduces photosynthetic efficiency because
of the Michaelis kinetics of CO fixation by the enzyme Rub-
isco (ribulose bisphosphate carboxylase oxygenase) (Fig.
1E). Calcification (Fig. 1C) provides an alternative proton
source and potentially allows autotrophs to avoid most of
the alkalinization and CO depletion that otherwise accom-
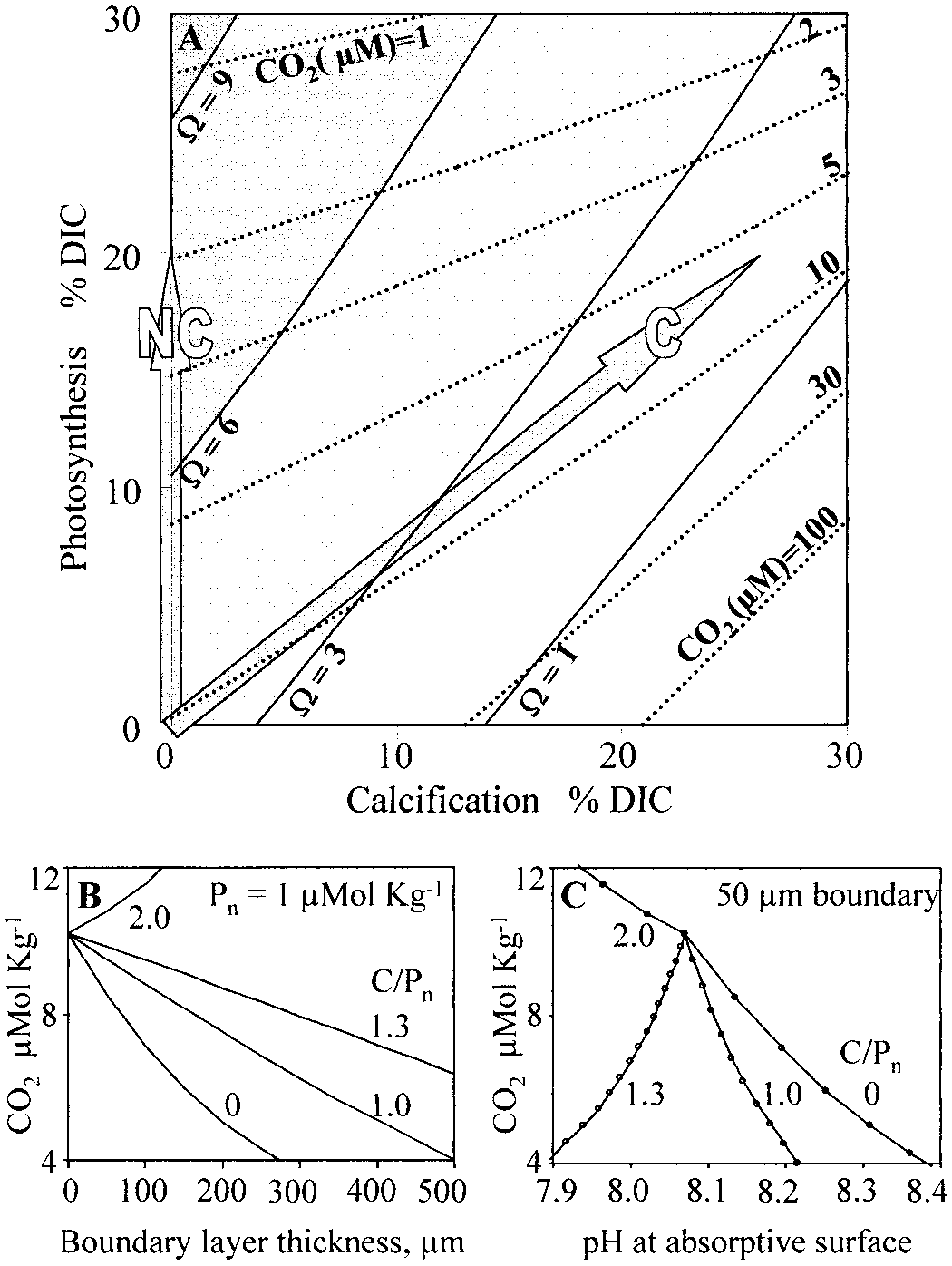
Figure 2A illustrates how photosynthesis and calcification
affect the aragonite saturation state (⍀) and molecular CO
(reaction 1) was originally postulated to account for 18O and
content of tropical surface seawater. Block arrows labeled
13C deficiencies in coral skeletons, which appeared to be
‘‘C’’ and ‘‘NC’’ represent hypothetical calcareous and non-
caused by kinetic discrimination against the heavy isotopes
calcareous autotrophs that photosynthetically remove 20% of
hydroxylation and hydration reactions (Mc-
the dissolved inorganic carbon (DIC) initially found in the
Connaughey 1989). The conversion of HCO Ϫ to CO (re-
seawater. The calcifier has a molar ratio of calcification to
action 2) is often catalyzed and can theoretically occur either
net photosynthesis (C : P ) of 1.3, which is about average for
within the extracellular boundary layer or intracellularly af-
reef corals (Gattuso et al. 1999). Although the calcifier and
ter importation of HCO Ϫ, Hϩ combinations. Alkaline and
noncalcifier have equal photosynthetic rates, the calcifier de-
acidic conditions favor reactions 1 and 2, respectively, and
pletes molecular CO about one-half as much—by 43%,
most calcareous organisms develop recognizable, highly al-
compared to the 81% rate associated with the noncalcifier.
kaline calcareous zones that contrast with the noncalcareous,
If both obey the same Michaelis curve for CO fixation (Fig.
absorptive regions in which HCO Ϫ assimilation occurs.
Carbon assimilation strategies. (A) CO uptake. (B)
HCO Ϫ assimilation using proton equivalents from solution. (C)
HCO Ϫ assimilation using protons from calcification. (D) Phos-
Photosynthesis, calcification, and seawater chemistry.
phate-proton cotransport. (E) Michaelis curve, illustrating kinetics
(A) Effects of photosynthesis (Y axis) and calcification (X axis) on
of CO fixation by the enzyme Rubisco. ‘‘C’’ and ‘‘NC’’ refer to
molecular CO concentrations (dotted lines, in Mol kgϪ1) and the
the hypothetical calcifier and noncalcifier, respectively, in Fig. 2A.
aragonite saturation state (⍀, solid lines with shading) of tropicalsurface seawater. Initial conditions at origin are described in meth-ods. Block arrows ‘‘C’’ and ‘‘NC’’ represent hypothetical calcareousand noncalcareous autotrophs, respectively. (B) CO concentrations
These are represented by the lower and upper surfaces, re-
at an autotroph’s absorptive surface for C : P ratios of 0, 1.0, 1.3,
and 2.0, plotted as a function of the boundary-layer thickness. Pn
To successfully couple reactions 1–4, the autotroph bal-
ϭ 1 Mol C mϪ2 sϪ1. (C) CO and pH at the autotroph’s absorptive
ances the photosynthetic benefits of higher CO concentra-
surface for a boundary-layer thickness of 50
tions (Fig. 1E) against the costs of calcification. Calcification
ϭ 1 Mol C mϪ2 sϪ1 are plotted as dots along the curves.
Connaughey and Whelan 1997). Each ‘‘CH O’’ unit pro-
duced during photosynthesis generates about 6 ATP upon
ences calcification rates by individual organisms that stim-
respiration, so the cost : benefit ratio of coupling calcification
ulate photosynthesis through calcification.
and gross photosynthesis is about (C ϫ 2)/(P ϫ 6) ϭ 1/3
C : P . C : P ratios above 3 appear to be energetically unpro-
Materials and methods—Experiments were carried out in
a coral reef microcosm located at the Marine Systems Lab-
As the protons from calcification are discharged into am-
oratory of the Smithsonian Institution of Natural History
bient waters (or the boundary layer adjacent to the absorp-
(Adey 1983; Small et al. 1998). The microcosm’s main tank
tive surface; Fig. 1C), not all react with HCO Ϫ to produce
held 400 liters, and associated algal turf scrubbers contained
CO . Some protons leak from the absorptive to the calcifying
an additional 1,280 liters. Natural sunshine supplied about
surface, and more importantly, some are soaked up by bases
one-third of the total light, and metal halide bulbs provided
such as OHϪ, CO ϭ, and B(OH) OϪ rather than HCO Ϫ. The
the remainder. Photosynthetic calcifiers included green algae,
alkalinity of solution, which measures the concentration of
coralline reds, foraminifera, stony corals, and giant clams.
all bases, therefore affects how much CO can be produced
Calcium and bicarbonate were replenished daily to maintain
through calcification. Conversely, acids in solution supple-
Ca2ϩ Ͼ 10.5 mM LϪ1 and total alkalinity Ͼ 2.40 mEq LϪ1
ment the protons from calcification. Community metabolism
during the period when measurements were made in the mi-
affects seawater alkalinity and acidity and potentially influ-
Changes in water chemistry during daytime incubations of algae and corals. (A) pH.
(B) Total alkalinity. (C) Calculated changes in CO and aragonite saturation states. (D) Coral ϩ
Chondria incubations. Dashed lines, microcosm; M ϭ Montipora, squares; A ϭ Acropora, triangles;C ϭ Chondria, circles; Ht and Ho ϭ Halimeda tuna (open diamonds) and H. opuntia (filled dia-monds), respectively. n ϭ 5 Replicates, except Chondria and H. tuna, for which n ϭ 2; standarddeviations are somewhat larger than markers.
For various experiments, corals and algae were placed in
cium concentrations were assigned seawater values (B ϭ
1- and 3-liter chambers that were suspended within the mi-
416 ϫ S/35; Ca2ϩ ϭ 10280 ϫ S/35 M kgϪ1) (Millero
crocosm. Water-filled chambers provided controls. Pumps
1995). Chemical modeling not tied to the microcosm as-
circulated water within the chambers. Incubations lasted 10
sumed that t ϭ 25ЊC and S ϭ 35‰, initially with 10.2 M
h, with sampling for O and pH every 2 h and sampling for
kgϪ1 dissolved molecular CO ; 1,962 M kgϪ1 total DIC;
alkalinity every 4 h. Five replicates were conducted for Ac-
2,300 Eq kgϪ1 alkalinity; and pH 8.0697. Millero’s (1995)
ropora, Montipora, and Halimeda tuna and for combinations
carbonate system thermodynamics were used throughout.
of Acropora or Montipora with Chondria. Three replicateswere conducted with Chondria and two were conducted for
Results—The noncalcareous alga Chondria sp. increased
pH but did not change alkalinity, whereas highly calcareous
Dissolved oxygen was measured by Winkler titrations,
species reduced in terms of alkalinity and did not elevate
with a precision based on two to three titrations per sample
pH as much (Fig. 3A,B). Chondria reduced molecular CO2
of Ϯ0.94 M LϪ1 (n ϭ 15). Titration alkalinity was deter-
concentrations by 73% and increased the aragonite saturation
mined with a precision for three replicates of Ϯ0.03 mEq
state from ⍀ ഠ3 to 6.2 (Fig. 3C). The corals Acropora and
LϪ1 (n ϭ 15). Prior to obtaining measurements, pH elec-
Montipora reduced CO by only 22 and 21%, respectively,
trodes were calibrated in Fisher-brand buffers (pH, 7.00 and
and reduced ⍀ by 11 and 17%, respectively. The lightly
10.00; NIST scale) with a precision of Ϯ0.02, and pH elec-
calcareous H. tuna behaved somewhat like the noncalcareous
trodes were later recalibrated to the seawater scale (Millero
Chondria, whereas the more calcareous H. opuntia behaved
1995) through measurements of a seawater carbon standard
provided by Andrew Dixon (Scripps Institution of Ocean-
Calcification and photosynthesis generally correlated over
ography). DIC, CO concentration, and aragonite saturation
the course of the day. Acropora and sometimes H. opuntia
state were calculated from pH and alkalinity data.
achieved ratios of calcification to net photosynthesis (C : P )
Calcification was estimated as one-half of alkalinity
as high as 1. Calcification and photosynthesis generally de-
change, and photosynthesis was estimated as the DIC change
clined during the afternoon, with photosynthesis declining
minus calcification (Smith and Kinsey 1978). Calculations
more sharply. The higher photosynthetic rates in the morning
based on microcosm experiments used observed tempera-
coincided with higher CO concentrations and lower pH and
tures (t ϳ 28ЊC) and salinities (S ϳ 36‰). Borate and cal-
O concentrations. Decreased calcification in the afternoon
occurred despite higher ⍀, which was, for example, 40%higher during the afternoon in the microcosm. Net calcifi-cation in the microcosm essentially ceased at night, eventhough the water remained supersaturated with respect toaragonite. Calcification and aragonite supersaturation werethus only loosely correlated. The noncalcareous Chondriaand lightly calcareous H. tuna raised ⍀ most strongly,whereas highly calcareous corals and H. opuntia left ⍀ un-changed (or even lowered ⍀) after several hours.
When Chondria and corals were incubated together, pH
increased as it did with Chondria alone, but alkalinitydropped rapidly. Calcification rates increased 60% (Acro-pora) and 130% (Montipora) compared with incubations ofcorals alone (Fig. 3D) (Student’s t-test, P ϭ 0.002 and P ϭ0.00002, respectively; n ϭ 5 in both cases.) Thus, the non-calcareous alga apparently stimulated calcification in nearbycorals. Photosynthetic rates in combined incubations werealso somewhat higher than were the sums from separate in-cubations, by values of 7% (Acropora) and 18% (Montipo-ra).
Projected ratio of calcification to photosynthesis (C : Pn
ϭ alkalinity : acidity; heavy line), aragonite saturation levels (⍀;
Discussion—Autotrophs balance benefits and costs in var-
solid line), and inorganic aragonite precipitation rate (dotted line,in
ious ways. Chondria and other fleshy algae reduce metabolic
mol CaCO mϪ2 sϪ1, calculated from kinetics of Burton and
Walter [1987]) in seawater as a function of pH (25ЊC, 35‰, con-
costs by not calcifying. (Photosynthesis by such organisms
stant total alkalinity ϭ 2,300 Eq kgϪ1).
alkalinizes the water and raises ⍀, but this does not causemuch calcification.) Corals and H. opuntia invest heavily incalcification and enjoy higher CO levels. The lightly cal-
cified H. tuna uses an intermediate strategy. The corals cal-
cify faster when incubated together with noncalcareous algae
McConnaughey and Whelan 1997). Even nonphotosynthetic
under more alkaline, CO -depleted conditions, indicating an
tissues can calcify through such physiologies. The ability of
active effort to forestall CO depletion.
calcifiers to locally elevate ⍀ well beyond ambient values
Ambient chemistry affects how much CO is generated by
contributes to the loose correlations between ambient ⍀ and
a particular amount of calcification. In reaction 4, HCO Ϫ
calcification rates observed in the present experiments.
supplies the autotroph with both carbon and protons, and C :
Inorganic aragonite precipitation from seawater obeys rate
P ratios ϭ 1. However, dissolved bases, including OHϪ,
equations such as R ϭ k(⍀ Ϫ 1)1.7 (Burton and Walter 1987)
B(OH) OϪ, and CO ϭ, compete with HCO Ϫ for protonation
(Fig. 4). The inorganic calcification rate doubles between pH
at the organism’s absorptive surface. This increases the
8.0 and 8.2, whereas the C : P ratio projected from the al-
amount of calcification required to produce CO (reaction 2)
kalinity : acidity ratio increases only 14%. (This assumes
by the factor [alkalinity]/[HCO Ϫ]. Dissolved acids, includ-
constant alkalinity. A constant DIC scenario produces sim-
ing CO and B(OH) , conversely supplement the protons
ilar results, whereas calcification increases more strongly
from calcification, reducing the amount of calcification re-
with pH under a constant CO scenario.) Calcification rates
quired by the factor [HCO Ϫ]/[acidity]. The C : P ϭ 1 ratio
predicted from ambient ⍀ and the alkalinity : acidity ratio are
Nutrients may also affect reef calcification rates. Highly
calcareous autotrophs such as corals (e.g., Marubini and Da-
Figure 4 plots this predictor of C : P ratio as a function of
vies 1996) and coccolithophorids (e.g., Paasche and Brubak
pH. Further adjustments related to the different diffusivities
1994) calcify faster when nutrients are scarce and thrive in
of various ions have only a small effect. Within the normal
nutrient-deficient waters. McConnaughey and Whelan
pH range of reef seawater (7.9–8.3), the C : P predictor lies
(1997) therefore suggested that calcification assists nutrient
between 1.0 and 1.3, a value that is similar to those values
uptake. Nutrient-proton cotransport (Fig. 1D) provides one
common among highly calcareous corals and algae. Sea-
likely mechanism. NO Ϫ and H PO Ϫ uptake in various stud-
water pH does not change when calcification and photosyn-
ies appear to involve cotransport of at least 1Hϩ and 2–4Hϩ,
thesis occur in a 1.2 ratio, which, according to Eq. 5, should
respectively (Sakano 1990; Wollenweber 1997). Energeti-
occur at a pH of 8.2. Higher pH raises the predicted C : P
cally, an ion entering a cell performs the chemical work RT
ratio and should stimulate calcification, nudging pH back
ln a /a , where R is the gas constant, T is Kelvin temperature,
down. This probably helps to maintain reef pH around 8.2.
and a and a are the activities of the chemical inside and
Calcifiers tend to isolate their calcification sites, as be-
outside of the cell. The electrical work is ZFV, where Z is
neath the aboral epithelium of corals, and then raise ⍀ lo-
the charge on the ion, F is the Faraday constant, and V is
cally through ion transport. pH often exceeds 10 at the cal-
the cell’s membrane potential. Adding these terms for a nu-
cification site of the alga Chara, and CO ϭ accumulates to
trient ion plus ‘‘n’’ protons and rearranging yields
log(a /a ) ϭ n(pH Ϫ pH ) ϩ (n ϩ Z)FV/2.3RTWalter H. Adey and Allegra M. Small
The final term accommodates additional energy terms such
as ATP hydrolysis. V and E are shown as positive so that
all terms are additive. If a cell increases (pH –pH ) by 0.5
pH units through calcification, it might therefore accumulate
NO Ϫ and H PO Ϫ at least three and 10–100 times more
strongly. Extracellular acidification should also improve
ADEY, W. H. 1983. The microcosm: A new tool for reef research.
NH ϩ uptake slightly and slow Fe2ϩ oxidation, aiding in as-
Coral Reefs 1: 193–201.
similation. Calcification’s potential for improving nutrient
BURTON, E. A., AND L. M. WALTER. 1987. Relative precipitation
assimilation therefore appears to be substantial. Moreover,
rates of aragonite and Mg Calcite from seawater: Temperature
seawater pH and alkalinity should affect this physiology
or carbonate ion control? Geology 15: 111–114.
much as they affect HCO Ϫ assimilation. Photosynthesis and
GATTUSO, J.-P., D. ALLEMAND, AND M. FRANKIGNOULLE. 1999.
calcification may therefore become correlated, as will cal-
Photosynthesis and calcification at cellular, organismal, and
cification and pH, even if the autotroph calcifies mainly to
community levels in coral reefs: A review of interactions and
control by carbonate chemistry. Am. Zool. 39: 160–183.
People increase nutrient inputs to many reefs through ag-
, S. REYNAUD-VAGANAY, I. BOURGE, P. FURLA, S. ROMAINE-
ricultural runoff, sewage disposal, increased erosion, and air-
LIOUD, M. FRANKIGNOULLE, AND J. JAUBERT. 2000. Calcifi-
borne nutrient transport. Nutrients may directly suppress cal-
cation does not stimulate photosynthesis in zooxanthellate scleractinian coral Stylophora pistillata. Limnol. Oceanogr. 45:
cification, as was observed, for example, by Marubini and
Davies (1996), and nutrients may encourage fleshy algae that
GOREAU, T. F. 1959. The physiology of skeleton formation in corals.
compete with the calcifiers and feed their predators. Through
Biol. Bull. 116: 59–75.
such mechanisms, nutrients may reduce reef calcification.
KAWAGUTI, S., AND D. SAKUMOTO. 1949. The effect of light on the
On the other hand, nutrients stimulate photosynthesis by
calcium deposition of corals. Bull. Oceanogr. Inst. Taiwan 4:
both calcareous and noncalcareous autotrophs. By raising pH
and the alkalinity : acidity ratio, reef calcification may be
MARUBINI, F., AND P. S. DAVIES. 1996. Nitrate increases zooxan-
stimulated, as was observed in the present coral ϩ Chondria
thellae population density and reduces skeletogenesis in corals.
experiments. This represents a metabolic cost to the calci-
Mar. Biol. 127: 319–328.
fiers and may reduce their competitiveness in nutrient-rich
MCCONNAUGHEY, T. A. 1989. 13C and 18O isotopic disequilibrium
situations. Nutrients probably have a greater potential for
in biological carbonates. II. In vitro simulation of kinetic iso-
affecting reef calcification rates than does anthropogenic
tope effects. Geochim. Cosmochim. Acta 53: 163–171.
CO , which will decrease the alkalinity : acidity ratio by only
, AND J. F. WHELAN. 1997. Calcification generates protons
10% as CO levels double. Industrial nitrogen fixation has
for nutrient and bicarbonate uptake. Earth-Sci. Rev. 42: 95–
already doubled the global rate of nitrogen fixation.
Returning to the introductory chicken-and-egg question,
MILLERO, F. J. 1995. Thermodynamics of the carbon dioxide system
calcification appears quite capable of stimulating photosyn-
in the oceans. Geochim. Cosmochim. Acta 59: 661–667.
thesis, but this process is not metabolically ‘‘free.’’ Photo-
PAASCHE, E., AND S. BRUBAK. 1994. Enhanced calcification in the
synthesis can also stimulate calcification. Photosynthesis in-
coccolithophorid Emiliania huxleyi (Haptophyceae) under
creases ambient ⍀, but this only slightly stimulates
phosphorus limitation. Phycologia 33: 324–330.
biological calcification. More importantly, photosynthesis in-
AKANO, K. 1990. Proton/phosphate stoichiometry in uptake of in-
organic phosphate by cultured cells of Catharanthus roseus
creases the alkalinity : acidity ratio, which reduces how ef-
(L.) G. Don. Plant Physiol. 93: 479–483.
ficiently calcification generates CO . More calcification is
SMALL, A. M., W. H. ADEY, AND D. M. SPOON. 1998. Are current
therefore needed to obtain a particular photosynthetic ben-
estimates of coral reef biodiversity too low? A view through
efit. Fleshy algae can thereby stimulate calcification in near-
the window of a microcosm. Atoll Res. Bull. 458: 1–20.
by corals. Coral calcification likewise counteracts CO de-
SMITH, S. V., AND D. W. KINSEY. 1978. Calcification and organic
pletion and may stimulate photosynthesis in nearby algae.
carbon metabolism as indicated by carbon dioxide, p. 469–484.
Nutrient levels meanwhile affect the competitiveness of cal-
In D. R. Stoddart and R. E. Johannes [eds.], Coral reefs: Re-
careous and noncalcareous autotrophs differently. Interac-
tions between calcifiers and noncalcifiers on reefs are thus
WOLLENWEBER, B. 1997. A sensitive computer-controlled pH-stat
system allows the study of net Hϩ fluxes related to nitrogen
uptake of intact plants in situ. Plant Cell Environ. 20: 400– 408.
1304 Cedar LaneSelah, Washington 98942-1717
1Corresponding author (coral2@wolfenet.com).
Parkinsonism and Related Disorders 14 (2008) 224–228Striatal FDOPA uptake and cognition in advanced non-dementedParkinson’s disease: A clinical and FDOPA-PET studyMarije van Beilena,b,Ã, Axel T. Portmana, Henk A.L. Kiersc, Ralph P. Maguirea,b,Valtteri Kaasinena, Marthe Koninga, Jan Pruimb,d, Klaus L. Leendersa,baDepartment of Neurology, University Medical Center Groningen, University
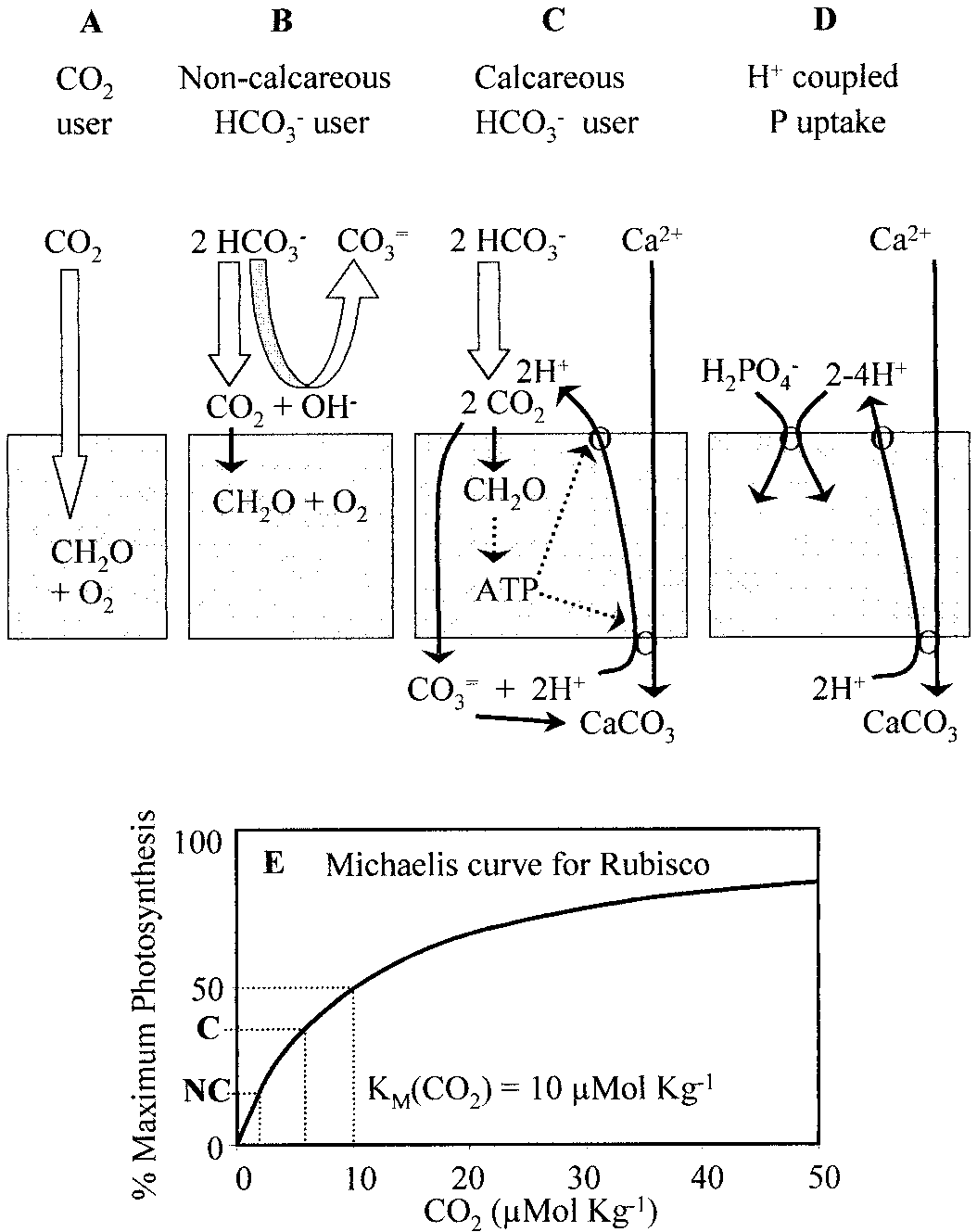
 Carbon assimilation strategies. (A) CO uptake. (B)
HCO Ϫ assimilation using proton equivalents from solution. (C)
HCO Ϫ assimilation using protons from calcification. (D) Phos-
Photosynthesis, calcification, and seawater chemistry.
Carbon assimilation strategies. (A) CO uptake. (B)
HCO Ϫ assimilation using proton equivalents from solution. (C)
HCO Ϫ assimilation using protons from calcification. (D) Phos-
Photosynthesis, calcification, and seawater chemistry.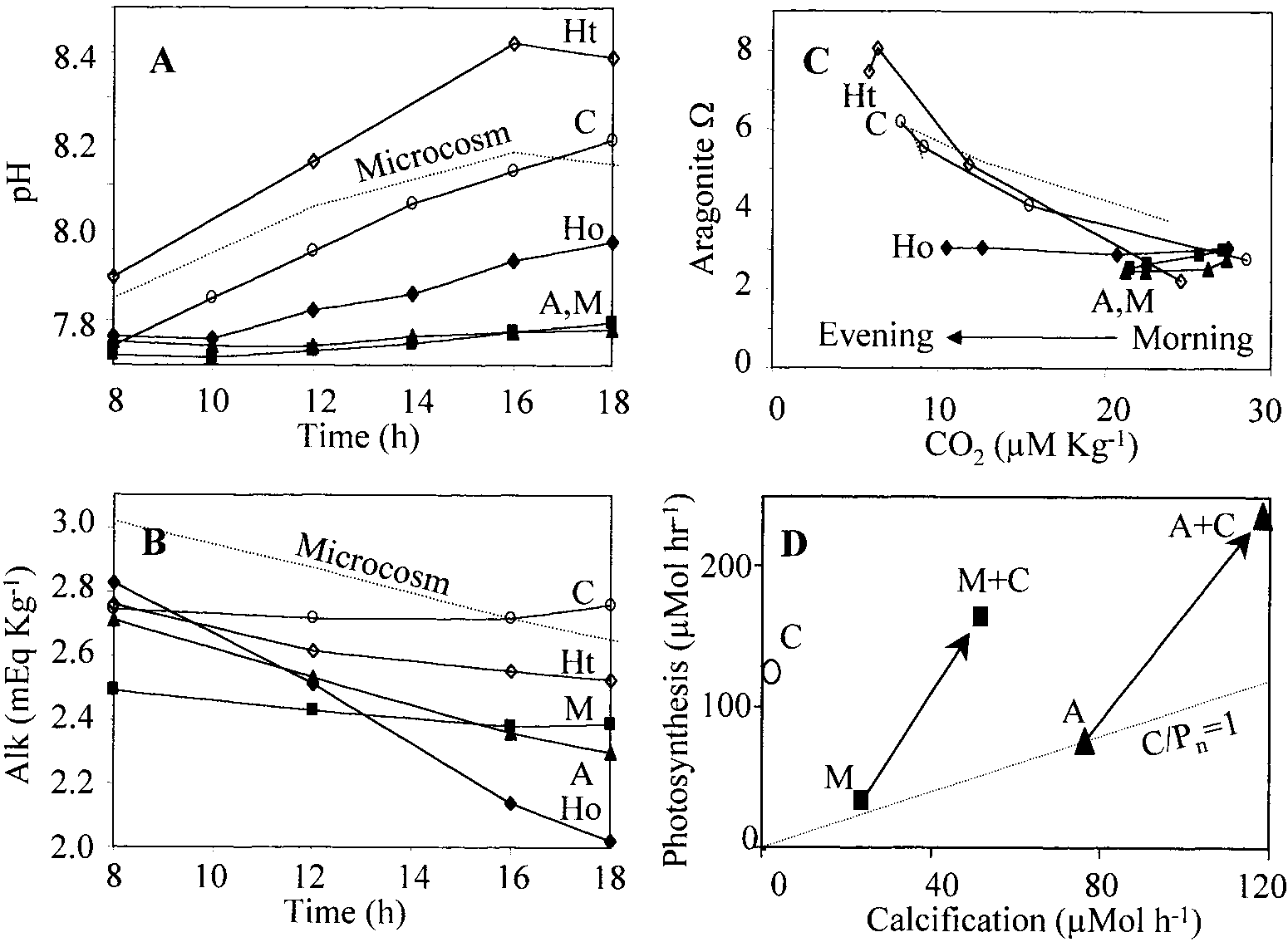 Changes in water chemistry during daytime incubations of algae and corals. (A) pH.
Changes in water chemistry during daytime incubations of algae and corals. (A) pH.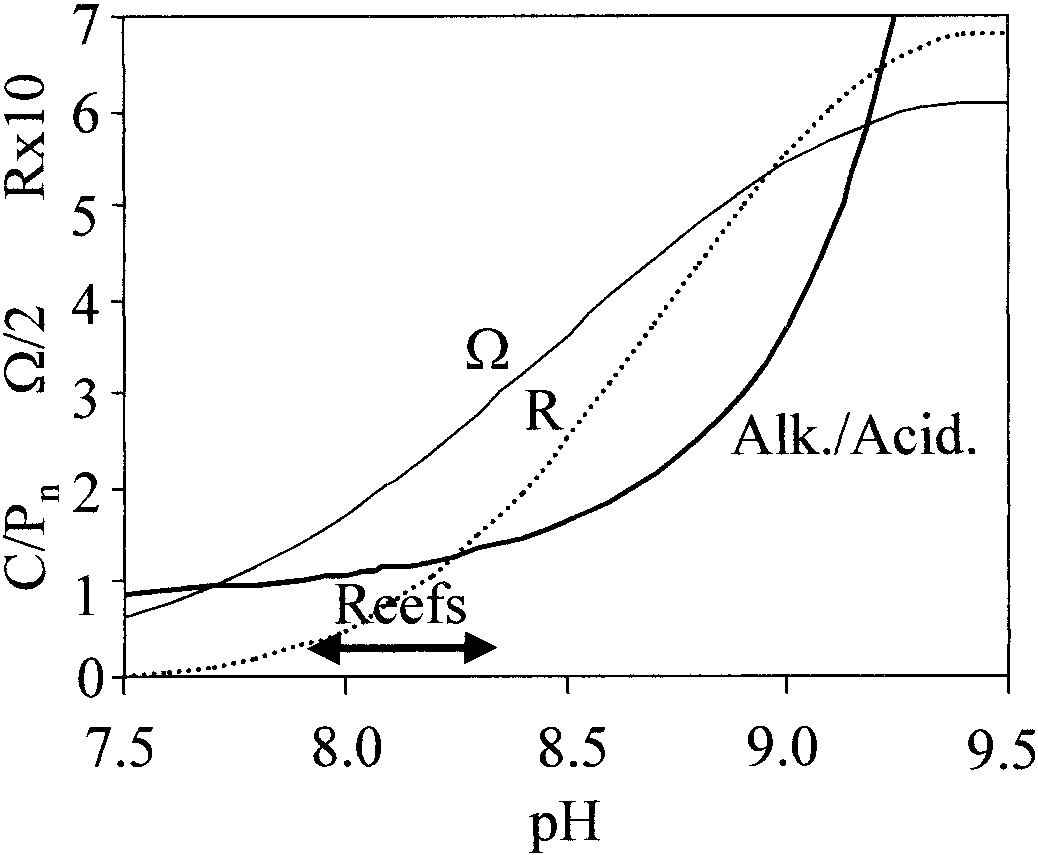 occurred despite higher ⍀, which was, for example, 40%higher during the afternoon in the microcosm. Net calcifi-cation in the microcosm essentially ceased at night, eventhough the water remained supersaturated with respect toaragonite. Calcification and aragonite supersaturation werethus only loosely correlated. The noncalcareous Chondriaand lightly calcareous H. tuna raised ⍀ most strongly,whereas highly calcareous corals and H. opuntia left ⍀ un-changed (or even lowered ⍀) after several hours.
occurred despite higher ⍀, which was, for example, 40%higher during the afternoon in the microcosm. Net calcifi-cation in the microcosm essentially ceased at night, eventhough the water remained supersaturated with respect toaragonite. Calcification and aragonite supersaturation werethus only loosely correlated. The noncalcareous Chondriaand lightly calcareous H. tuna raised ⍀ most strongly,whereas highly calcareous corals and H. opuntia left ⍀ un-changed (or even lowered ⍀) after several hours.