Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Microsoft word - art. 8 embriogenesis.doc
Ra Ximhai
Revista de Sociedad, Cultura y Desarrollo
Ra Ximhai Universidad Autónoma Indígena de México ISSN: 1665-0441 México
EMBRIOGÉNESIS SOMÁTICA EN (Anthurium andraeanum Lind.) VARIEDAD ‘LAMBADA’
Nydia del Rivero Bautista, Daniel Agramonte Peñalver, Raúl Barbón Rodríguez, Wilder
Camacho Chiu, Raúl Collado López, Felipe Jiménez Terry, Marta Pérez Peralta y Odalys
Ra Ximhai, enero-abril, año/Vol.4, Número 1
Universidad Autónoma Indígena de México
Mochicahui, El Fuerte, Sinaloa. pp. 135-149
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
EMBRIOGÉNESIS SOMÁTICA EN (Anthurium andraeanum Lind.) VARIEDAD ‘LAMBADA’ SOMATIC EMBRYOGENESIS IN (Anthurium andraeanum Lind.) VARIETY ‘LAMBADA’
Nydia del Rivero-Bautista1, Daniel Agramonte-Peñalver2, Raúl Barbón-Rodríguez2, Wilder Camacho-Chiu3, Raúl Collado-López2, Felipe Jiménez-Terry2, Marta Pérez- Peralta2 y Odalys Gutiérrez-Martínez2
1Colegio de Postgraduados Campus-Tabasco. Periférico Carlos A. Molina s/n. H. Cárdenas, Tabasco. CP 86500. Fax: (973) 3722297 *Autor para correspondência E-mail: delriverobautista@yahoo.com.mx; 2Instituto de Biotecnología de las Plantas. Universidad Central “Marta Abreu” de Las Villas. Carretera a Camajuaní km 5.5, Santa Clara, Villa Clara, Cuba. CP 54830. Fax: 53(42)281329. E-mail: agramonte@ibp.co.cu 3Dirección General Tecnológica Agropecuaria. Carretera Cumuapa, Cunduacán, Tabasco, México. E-mail: wildercamachochiu@hotmail.com
Varias concentraciones y tipos de reguladores de crecimiento fueron probados para inducir la embriogénesis somática indirecta en Anthurium andraeanum Lind. (Monocotiledónea). Para la formación de callos, se utilizaron como explantes segmentos foliares de plantas micropropagadas sobre un medio de cultivo MS modificado suplementado con 6,79 µM de ácido 2,4-diclorofenoxiacético (2,4-D) y 2,32 µM kin (kinetina). La diferenciación de los embriones somáticos se logró en el medio de cultivo MS modificado que contenía 4,44 µM de 6-bencilaminopurina (6-BAP), en condiciones de oscuridad. La germinación de los embriones somáticos se obtuvo en el mismo medio de cultivo probado, suplementado con 1,78 µM de 6-BAP cuando se colocaron en una cámara de crecimiento con luz solar. Palabras clave: Embriones somáticos, medios de cultivo, reguladores de crecimiento.
Several concentrations and types of growth regulators were tested to induce the indirect somatic embryogenesis in Anthurium andraeanum Lind (Monocot). For the callus formation, they were used as explants foliar segments of micropropagated plants on a culture medium MS modified supplemented with 6,79 µM of 2,4-dichlorophenoxyacetic acid (2,4-D) and 2,32 µM kin (kinetin). The differentiation of the somatic embryos was achieved in the culture medium MS modified containing 4,44 µM of 6-benzylaminopurine (6-BAP) under conditions of darkness. The germination of the somatic embryos was obtained in the same culture medium tested supplemented with 1,78 µM 6-BAP when placed in the sunlight growth room. Key words: Culture medium, growth regulators, somatic embryos.
Recibido: 10 de diciembre de 2007. Aceptado: 11 de febrero de 2008. Publicado como ARTÍCULO CIENTÍFICO en Ra Ximhai 4 (1): 135-149.
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
INTRODUCCIÓN
El Anthurium es el género más grande de las Araceae con 1000 especies (Croat, 1990).
Especies e híbridos dentro de este género son altamente apreciados como ornamentales por
sus hermosas flores y exótico follaje (Kuenhle et al., 1992). El Anthurium ha sido
propagado tradicionalmente por vástagos y mediante el cultivo in Vitro de yemas axilares y
callos, seguido por la formación de yemas adventicias; sin embargo este método resulta en
la formación de plantas fuera de tipo (Geier, 1990; Martin et al., 2003; Puchooa, 2005).
Métodos eficientes para la regeneración de plantas de células cultivadas de muchas especies
importantes de cultivo fueron desarrollados durante la etapa de los años 1980 del siglo XX.
Esto fue posible por la inducción de cultivos embriogénicos los cuales formaron embriones
somáticos que germinaron para dar lugar a plantas normales. Esta tecnología, combinada
con la habilidad de células vegetales genéticamente transformadas forman hoy las bases de
la biotecnología vegetal a nivel comercial (Vasil, 2003).
La embriogénesis somática es una herramienta útil para la propagación masiva y programas
de transformación genética para el Anthurium (Geier, 1982; Kuenhle et al., 1992; Hamidah
et al., 1997). La formación de los embriones somáticos es muy sensible a las condiciones
de cultivo tales como la composición del medio de cultivo, el ambiente físico de cultivo, el
genotipo y la fuente del explante (Fuentes et al., 2000). Zaidi et al., (2000) mencionan que
el medio de cultivo más utilizado para la embriogénesis somática en monocotiledóneas es
el MS (Murashige y Skoog, 1962) y en menor proporción el medio de cultivo NN (Nitsch y
Nitsch, 1969). Sin embargo, en el género Anthurium se han empleado los dos medios de
cultivo mencionados con respuestas diferentes por Pierik (1976); Geier (1986); Kuehnle et al., (1992); Atta-Alla et al., (1998) y Puchooa (2005).
El objetivo de esta investigación fue lograr la inducción de la embriogénesis somática
indirecta en Anthurium andraeanum Lind., variedad ‘Lambada’.
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
MATERIALES Y MÉTODOS Material vegetal
La presente investigación se realizó en el laboratorio de Propagación Masiva del Instituto
de Biotecnología de las Plantas perteneciente a la Universidad Central “Marta Abreu” de
Las Villas, ubicado en Santa Clara, Cuba.
Para iniciar los trabajos de embriogénesis somática se emplearon plántulas propagadas a
través de organogénesis indirecta según la metodología propuesta por Pierik (1976). Como
explantes iniciales se utilizaron hojas de plantas obtenidas in vitro de anturio variedad
‘Lambada’, a las cuales se les eliminaron los bordes y se seccionaron con un tamaño
aproximado de un centímetro cuadrado y estos se colocaron con la parte adaxial hacia el
El pH de los medios de cultivo fue ajustado a 5.8 antes de su esterilización y gelificados
con 2.5 g.l-1 de Gelrite (Sigma, MO, EE.UU).
Inducción y formación de callos
Para la inducción y formación de los callos se empleó el medio de cultivo propuesto por
Murashige y Skoog (1962) (MS) modificado con una reducción de los nitratos,
componentes de los macronutrientes a 50%, micronutrientes completos, vitaminas MS,
mio-inositol 100 mg.l-1, tiamina 0.4 mg.l-1, sacarosa 30 g.l-1. Suplementado con cuatro
concentraciones de 2,4-D (2.26; 4.52; 6.79 y 9.05 µM) combinadas con 2.32 µM de
Se utilizaron diez frascos de cultivo con cinco explantes para cada tratamiento. Las
condiciones de cultivo fueron de oscuridad y una temperatura de 27±2ºC. Se realizaron
observaciones cada ocho días para describir los cambios y características morfológicas de
los callos. A los 30 días de cultivo se evaluó el porcentaje de explantes que formaron callo.
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
Diferenciación de los embriones somáticos
La diferenciación de los embriones somáticos se desarrolló en el medio de cultivo
mencionado arriba y suplementado con tres concentraciones de 6-BAP (2.22; 4.44 y 8.88
µM) y un control sin reguladores de crecimiento.
Se emplearon diez frascos de cultivo con cinco callos como explantes para cada
tratamiento. Las condiciones de cultivo fueron de oscuridad y una temperatura de 27±2ºC.
A los 60 días de cultivo se observó la presencia de embriogénesis somática de alta
frecuencia (ESAF) y embriogénesis somática de baja frecuencia (ESBF). Las variables
evaluadas fueron el número de callos con embriones somáticos, el número de embriones
somáticos por callo y el número de embriones somáticos que alcanzaron la etapa
Germinación de los embriones somáticos
La germinación de los embriones somáticos ocurrió en un medio de cultivo MS modificado
donde se estudiaron tres concentraciones de 6-BAP (0.89, 1.78 y 2,66 µM), con una
temperatura de 25±2ºC y una densidad luminosa de flujo de fotones fotosintéticos (FFF) de
100-125 µmol.m-2.s-1. Se emplearon seis embriones somáticos en etapa coleoptilar por
frasco de cultivo y se utilizaron 24 frascos como repeticiones.
Se efectuaron observaciones diarias para observar el inicio de la germinación y a los 30
días de cultivo se evaluó el número de embriones somáticos con germinación completa,
número de embriones somáticos con germinación parcial y número de embriones somáticos
con embriogénesis somática secundaria.
Diseño experimental y análisis estadístico
Los experimentos fueron desarrollados con un diseño completamente al azar. A los datos
obtenidos se les realizaron la comprobación de los supuestos de normalidad y
homogeneidad de varianza. Los datos experimentales fueron analizados con un análisis de
varianza simple. La comparación de medias en los tratamientos se efectuó mediante la
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
prueba de rangos múltiples según Tukey y Dunnett’s con una significación de 0.05,
mediante el programa estadístico SPSS versión 13.0 para ‘Windows’.
RESULTADOS Y DISCUSIÓN Inducción y formación de callos
La formación de callos en los explantes comenzó en las zonas de corte de los segmentos
foliares en todos los tratamientos evaluados. Los callos formados cubrieron la totalidad del
segmento foliar a los 45 días de cultivo, donde se observó la presencia de un callo
compacto, de color amarillo translúcido con características nodulares (Figura 1).
Figura 1. Callos formados a partir de segmentos foliares en Anthurium andraeanum Lind. variedad
‘Lambada’ en el medio de cultivo MS modificado a los 45 días de cultivo.
El mejor tratamiento empleado para la inducción y formación de callos fue donde se utilizó
una concentración de 6.79 µM de 2.4-D combinado con 2.32 µM de kinetina, en este se
obtuvo el mayor porcentaje de explantes con formación de callos (Cuadro 1).
Cuadro 1. Efecto de la concentración de 2,4-D en la inducción y formación de callos a partir de segmentos foliares de Anthurium andraeanum variedad ‘Lambada’ a los 45 días de cultivo. Concentración de Número de Porcentaje de regulador de explantes con explantes con crecimiento (2,4-D) formación de formación de callos
Medias con letras distintas en una misma fila difieren para p< 0.05 según prueba de Dunnett’s . EE=error estándar.
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
En cuanto al efecto del 2,4-D en la formación de callos; resultados similares fueron
encontrados por Kuehnle et al., (1992) y Matsumoto y Kuehnle (1996) en híbridos de A. andraeanum, los cuales alcanzaron la inducción y formación de callos en los explantes con
un rango que varió desde 4.52 a 18.1 µM de 2,4-D y 1.39 a 4.56 µM de kinetina en un
medio de cultivo MS modificado. Por el contrario, en esta misma especie Kuehnle y Suggii
(1991) y Puchooa (2005), indujeron una mayor formación de callos con una combinación
de 0.45 µM 2,4-D y 4.44 µM de BA. Otros autores como Hamidah et al., (1997) y Geier
(1986) en A. scherzerianum lograron la formación de callos (61.4%) con 6.75 µM 2,4-D
combinado con 2.3 µM kinetina en un híbrido y con 0.36 µM 2,4-D suplementado con 4.4
µM BA, en un medio de cultivo Nitsch y Nitsch (1969) modificado en 18 genotipos,
respectivamente. En otra monocotiledónea, como Hevea brasiliensis (Muell.) Arg., la
inducción y formación óptima de callo se alcanzó en un medio de cultivo MS modificado
suplementado con 9.05 µM de 2,4-D y 2.32 µM de kinetina (Kumari et al., 1999).
Nakano et al., (2004) en especies del género Tricyrtis una planta ornamental, obtuvieron la
mayor formación de callo con estructuras embriogénicas con 4.5 µM de 2,4-D y 0.045 µM
de TDZ. En otras especies como trigo (Triticum sp.) y caña de azúcar (Saccharum officinarum L.), Zale et al., (2004) y Gandonou et al., (2005) indicaron que la habilidad en
la inducción de callos está fuertemente influenciada por el genotipo.
Diferenciación de los embriones somáticos
En los callos empleados como explantes y colocados en el medio de cultivo para la
formación y diferenciación de los embriones somáticos, a partir de los 40 días de cultivo se
observó la presencia de pequeñas estructuras de coloración amarillo transparente en los
callos a los 60 días de cultivo, además en un gran número de callos se observó la presencia
de ESAF y ESBF. La embriogénesis somática de alta frecuencia se caracterizó por la
presencia de grupos de proembriones y también de embriones somáticos en etapa globular
agrupados de color amarillo transparente; sin embargo, en la embriogénesis somática de
baja frecuencia se observaron embriones somáticos aislados en diferentes etapas de
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
Figura 2. (A)- Callo con embriogénesis de alta frecuencia (ESAF) obtenido a partir de explantes foliares de Anthurium andreanum con 4,44 µM de 6-BAP a los 40 días de cultivo, (B)- Callo con embriogénesis de baja frecuencia (ESBF).
En cuanto al número de callos con embriones somáticos el mayor porcentaje (68,0%) se
presentó en el tratamiento que contenía 4.44 µM de 6-BAP con diferencias significativas
con el resto de los tratamientos. La presencia de los embriones somáticos en los callos
cultivados con esta citoquinina demostró el efecto de la misma en la formación de
embriones somáticos en anturio variedad ‘Lambada’ (Cuadro 2).
En lo que respecta al número de embriones somáticos por callo los valores más altos se
obtuvieron en un medio de cultivo con 8.88 µM de 6-BAP, con diferencias con respecto al
resto de los tratamientos (Cuadro 2). A medida que se aumentó la concentración de 6-BAP
se incrementó la formación de embriones somáticos por callo. Sin embargo, el mayor
porcentaje (84.2%) de embriones somáticos que alcanzaron la etapa coleoptilar se logró con
Cuadro 2. Efecto de diferentes concentraciones de 6-BAP en el desarrollo de los embriones somáticos en medio de cultivo MS modificado en anturio (A. andraeanum Lind.) a los 80 días de cultivo. Concentraciones Número de Porcentaje *Número de Número de Porcentaje de callos con de callos embriones embriones embriones embriones somáticos somáticos somáticos en somáticos embriones por callo somáticos coleoptilar coleoptilar
Medias con letras diferentes en una misma columna difieren según la prueba por Tukey para p<0.05. *Medias con letras diferentes en la misma fila difieren según la prueba por Dunnett’s para p<0.05. EE= Error estándar.
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
Con respecto a los resultados alcanzados estos difieren con los encontrados por Kuehnle et al., (1992) quiénes inducen la formación de embriones somáticos en A. andraeanum con
6.79 µM de 2,4-D combinado con 2.32 µM de kinetina y con Hamidah et al., (1997)
quienes indujeron la producción de embriones somáticos con 0.46 µM de kinetina en A. scherzerianum. Sin embargo, concuerdan con Shibli y Ajlouni (2000) en black iris (Iris nigricans) una especie ornamental que con una cantidad similar 4.5 µM de BA lograron el
mayor número de embriones somáticos por callo. Otros autores, como Nakano et al.,
(2004) en Liliaceae hallaron la formación de embriones somáticos en un medio de cultivo
libre de reguladores de crecimiento. Choi et al., (1998) mencionan que el desarrollo de
embriones en tejido somático se logró en ausencia de reguladores de crecimiento así como
también con la presencia de otros reguladores de crecimiento. Se ha demostrado que las
citoquininas son necesarias para el desarrollo de los embriones somáticos (Fujimura y
Komamine, 1980; Tan et al., 2001). En caucho (Hevea brasiliensis) Kumari et al., (1999) el
máximo número de embriones somáticos se desarrolló en un medio de cultivo
suplementado con 3.25 µM de kinetina y 1.07 µM de ANA.
Por otra parte, Vega y Prehn (2005) en Quillay y Lee y Lee (2003) en Dicentra spectabilis
al suplementar el medio de cultivo con 6-BAP hallaron un incremento en la diferenciación
de los embriones somáticos, demostrando el efecto de las citoquininas. En el cultivo de
rosas en la variedad ‘Carefree Beauty’ y en Musa con la adición de 2.2 µM de 6-BAP al
medio de cultivo se alcanzó la mayor frecuencia en la diferenciación de los embriones
somáticos (Khalil et al., 2002; Li et al., 2002). George (1993) señala que las citoquininas
promueven el crecimiento de los embriones preformados cuando son adicionados al medio
de cultivo de formación de embriones somáticos.
Germinación de los embriones somáticos
Cuando se evaluó el efecto de diferentes concentraciones de 6-BAP en la germinación de
embriones somáticos en Anthurium andraeanum Lind., se pudo observar que el mayor
porcentaje de embriones somáticos con germinación completa y parcial se obtuvo en el
tratamiento con 1,78 µM de 6-BAP con diferencias significativas con el resto de los
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
tratamientos (Cuadro 3). Los valores de esta variable disminuyeron con el incremento de
las concentraciones de 6-BAP y con el control.
Cuadro 3. Efecto de diferentes concentraciones de 6-BAP en la germinación de embriones somáticos en Anthurium andraeanum Lind., a los 60 días de cultivo. Tratamientos Germinación completa de los Germinación parcial de los embriones somáticos embriones somáticos No. embriones No. embriones somáticos/frasco somáticos/frasco Media Porcentaje *Rangos Media Porcentaje
Medias con letras desiguales en una misma columna difieren según la prueba de Tukey para p<0.05. *Rangos medios con letras desiguales en una misma columna difieren según la prueba de Kruskal-Wallis para p<0.05. EE= Error estándar
Los resultados alcanzados difieren con los obtenidos por Kuehnle et al., (1992) quienes con
una concentración de 0.89 µM de 6-BAP lograron la germinación de embriones somáticos
de híbridos de A. andraeanum. Algunos autores han señalado que la adición de 6-BAP al
medio de cultivo incrementa la germinación de los embriones somáticos (Kuehnle et al.,
1992; Laxmi et al., 1999; Ashok et al., 2002; Barbón et al., 2002; Li et al., 2002). Sin
embargo, Mohammed et al., (2002) y Nakano et al., (2004) en estatice (Limonium bellidifolium) y lirios (Tricyrtis spp.) encontraron que los embriones somáticos en etapa
cotiledonal germinaron después de dos a tres días sobre medio basal MS carente de
En todos los tratamientos se observaron embriones somáticos con embriogénesis somática
secundaria, pero el mayor porcentaje (53.33%) de esta variable se alcanzó cuando se le
adicionó al medio de cultivo 2.66 µM de 6-BAP con diferencias significativas con el resto
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
Tabla 4. Influencia de diferentes concentraciones de 6-BAP en el número de embriones somáticos que desarrollaron embriogénesis somática secundaria en Anthurium andraeanum variedad ‘Lambada’ a los 60 días de cultivo. Tratamientos Embriones somáticos con embriogénesis secundaria No. de ES por frasco Rangos medios Porcentaje
Rangos medios con letras distintas en una misma columna difieren según prueba de Kruskal-Wallis para p<0.05.
Ravishankar y McComb (2006) encontraron en sándalo (Santalum album L.) que la
producción de embriones somáticos secundarios a partir de los embriones primarios ocurrió
cuando los embriones somáticos se mantuvieron por períodos largos de tiempo sobre
medios de cultivo de inducción con 6-BAP o TDZ. La formación de embriogénesis
secundaria es una de las causas de la asincronía durante la embriogénesis somática, los
embriones somáticos pueden formarse a partir del eje principal del embrión (Gavich et al.,
1992) y esto es debido a que existe una inducción de la división de las células
embriogénicas que quedan latentes en las paredes del embrión (Nomura y Komamine,
De los resultados presentados en este estudio permiten concluir que la combinación de 6.79
µM de 2,4-D y 2.32 µM de kinetina en un medio de cultivo MS modificado fue donde se
alcanzaron el mayor número de explantes con formación de callos. La diferenciación de los
embriones somáticos se logró en el mismo medio de cultivo suplementado con 4.44 µM de
6-BAP. Por último la germinación de los embriones somáticos se obtuvo con una
AGRADECIMIENTOS
Nuestro agradecimiento a la Asociación Nacional de Universidades e Instituciones de
Educación Superior (ANUIES) y al Colegio de Postgraduados Campus Tabasco por la beca
y el apoyo otorgado para la realización de esta investigación y los estudios de Doctorado.
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
LITERATURA CITADA
Ashok, K. H.G., Murthy, H. N. y Paek, K. Y. 2002. Somatic embryogenesis and plant regeneration in Gymnema silvestre. Plant Cell, Tissue and Organ Culture 71:85–88.
Atta-Alla, H., McAlister, B. G. y Van, S. J. 1998. In vitro culture and establishment of Anthurium parvispathum. S. Afr. J. Bot 64(5):296-298.
Barbón, R., Jiménez, E., De Feria, M., Quiala, E., Capote, A. y Chávez, M. 2002.
Influencia del genotipo y la densidad de inoculación sobre la diferenciación de embriones somáticos de Coffea arabica L. cv. Caturra rojo y Coffea canephora cv. Robusta. Biotecnología Vegetal 3:145-148.
Choi, Y. E., Yang, D. C., Park, J. C., Soh, W. Y. y Choi, K. T. 1998. Regenerative ability of somatic single and multiple embryos from cotyledons of Korean ginseng on hormone-free medium. Plant Cell Rep. 17:544.551.
Croat, T. B. 1990. A comparison of ardid classification systems.Aroideana 13:44-63.
Fuentes, R.L.S., Calheiros, B. P. M., Manetti-Filho J. y Vieira, G. E. L. 2000. The effects of silver nitrate and different carbohydrate sources on somatic embryogenesis in Coffea canephora. Plant Cell, Tissue and Organ Culture 60:5-13.
Fujimura, T. y Komamine, A., 1980. The serial observation of embryogenesis in a carrot cell suspension culture. New Phytol. 86:213-218.
Gandonou, Ch., Errabii, T., Abrini, J., Idaomar, M., Chibi, F., Senhaji, N.S. 2005. Effect of genotype on callus induction and plant regeneration from leaf explants of sugarcane (Saccharum sp.). African Journal of Biotechnology 4(11):1250-1255.
Gavich, H., Vardi, A., Fluhr, R. 1992. Suppression of somatic embryogenesis in Citrus cell cultures by extracellular proteins. Planta 186:511-517.
Geier, T. 1982. Morphogenesis and plant regeneration from spadix fragments of Anthurium scherzerianum cultivated in vitro. In A. Fujiwara (ed.) Proceeding of
the Fifth International Congress Plant Tissue and Cell Culture, 137-138. Japanese
Association for Plant Tissue Culture, Japan.
Geier, T. 1986. Factors affecting plant regeneration from leaf segments of Anthurium scherzerianum Schott (Araceae) cultured in vitro. Plant Cell Tissue and Organ
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
Geier, T. 1990. Anthurium. En: Ammirato, PV, Evans DA, Sharp WR y Bajaj YPS (eds.).
Handbook of plant cell and tissue culture. Vol. 5. McGraw-Hill, New York. P. 228-
George, E. E. 1993. Plant micropropagation of tissue culture:sugars-nutritional and regulatory effects. London:Exegeties, 322-336.
Hamidah, M., Ghani-Abdul, K. A. G. y Debergh, P. 1997. Somatic embryogenesis and planta regeneration in Anthurium scherzerianum. Plant Cell, Tissue and Organ
Khalil, S. M., Cheah, K. T., Perez, E. A., Gaskill, D. A. y Hu, J. S. 2002. Regeneration of banana (Musa spp. AAB cv. Dwarf Brazilian) via secondary somatic
embryogenesis. Plant Cell Rep 20:1128–1134.
Kuehnle, A. R. y Sugii, N. 1991. Callos induction and plantlet regeneration in tissue cultures of Hawaiian Anthuriums. HortSci 26:919-921.
Kuehnle, R. A., Chen, F. Ch. y Sugii, N. 1992. Somatic embryogenesis and plant regeneration in Anthurium andraeanum hybrids. Plant Cell Reports 11:438-442
Kumari, J. P., Asokan, P. M., Sobha, S., Sankari, A. L., Rekha, K., Kala, G. R., Jayasree, R.
y Thulaseedharan, A. 1999. Somatic embryogenesis and plant regeneration from immature anthers of Hevea brasiliensis (Muell.). Arg. Curr. Sci. 76:1242–1245.
Laxmi, D. V., Sharma, H. C., Kirti, P. B. y Mohan, M. L. 1999. Somatic embryogenesis in mango(Mangifera indica L.) cv. Amrapali. Current Science 77(10):1355-1358.
Lee, K. P., y Lee, D. W. 2003. Somatic embryogenesis and plant regeneration from seeds of wild Dicentra spectabilis (L.) LEM. Plant Cell Reports 22:105-109.
Li, X., Krasnyanski, F. S. y Korban, S. S. 2002. Somatic embryogenesis, secondary somatic embryogenesis, and shoot organogenesis in Rosa. J. Plant Physiol.
Martin, K. P., Dominic, J., Madassery, J. y Phillip, V. J. 2003. Direct shoot regeneration from lamina explants of two commercial cut flower cultivars of Anthurium andraeanum Hort. In Vitro Cell. Dev. Biol.-Plant 39:500-504.
Matsumoto, T. K. y Kuenhle, A. R. 1996. Anthurium micropropagation. En: Bajaj YPS
(Ed). Biotechnology in agriculture and forestry: High technology and
micropropagation. Springer Verlag, New York. 40:14-29.
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
Mohammed, A. M. A., Rathinasabapathi, B. y Kelley, K. 2002. Somatic embryogenesis in perennial statice Limonium bellidifolium, (Plumbaginaceae). Plant Cell, Tiss.
Murashige, T. y Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plantarum 15: 473-497.
Nakano, M., Mizunashi, K., Tanaka, S., Godo, T. T., Nakata, M. y Saito, H. 2004.
Somatic embryogenesis and plant regeneration from callus culture of several species in the genus Tricyrtis. In Vitro Cell. Dev. Biol.-Plant 40:274–278.
Nitsch, J. y Nitsch, C. 1969. Haploid plants from pollen grains. Science 163:85-87.
Nomura, K. y Komamine, A. 1995. Physiological and biochemical aspects of somatic embryogenesis. En: Thorpe, T.A. (ed). In vitro embryogenesis in plants. Kluwer
Pierik, R. L. M. 1976. Anthurium andraeanum plantlets produced from callus tissues cultivated in vitro. Physiol. Plant. 37:80-82.
Puchooa, D. 2005.In vitro mutation breeding of Anthurium by gamma radiation. Int.
Ravishankar y McComb. 2006. Factors influencing in vivo and in vitro micrografting of sandalwood (Santalum album L.): an endangered tree species. In: Journal of
Forest Reasearch. núm. 3, año/vol. 11. págs. 147-151.
Shibli, A. R. y Ajlouni, M. M. 2000. Somatic embryogenesis in the endemic black iris.
Plant Cell, Tissue and Organ Culture 61:15-21.
Tan, N. D., van, Le Bui y Thanh, V. T. 2001. Manipulation of the morphogenetic pathways of Lilium longiflorum transverse thin cell layer explants by auxin and cytokinin. In Vitro Cell. Dev. Biol.-Plant 37:44-49.
Vasil, K. I. 2003. Somatic embryogenesis and its applications to plant biotechnology. V
Reunión de la Sociedad Española de Cultivo in vitro de Tejidos Vegetales. 29 Junio-2
Vega, A. y Prehn, D. 2005. Inducción e inicio de maduración in vitro de tejido embriogénico de Quillaja saponaria. Ciencia e Investigación Agraria 32(3):197-
Embriogénesis somática en (Anthurium andraeanum Lind.) variedad ‘LAMBADA’
Zaidi, N., Habib, N. K., Zafar, F. y Zafar, S. 2000. Bulbous and cormous monocotyledonous ornamental plants in vitro. Quarterly Science Vision 6(1):58-
Zale, J. M., Borchardt-Wier, H., Kidwell, K. K. y Steber, C. M. 2004. Callus induction and plant regeneration from mature embryos of a diverse set of wheat genotypes. Plant Cell, Tissue and Organ Culture 76:277-281. Nydia del Rivero Bautista Bióloga por el Colegio de Postgraduados Campus-Tabasco. Daniel Agramonte Peñalver Doctor en Ciencias en Universidad Central Marta Abreu de las Villas, Instituto de Biotecnología de las Plantas, Santa Clara, Cuba. Maestro en Ciencias en Biotecnología Vegetal, Instituto de Biotecnología Vegetal. Santa Clara, Cuba. Licenciatura en la Universidad Central Marta Abreu de las Villas, Facultad de Ciencias Agropecuarias. Santa Clara Cuba. Profesor Investigador del Instituto de Biotecnología de las Plantas desde 1994. Director del Institución de Biotecnología de las Plantas desde 2002. Raúl Barbón Rodríguez Doctor y Profesor Investigador Titular por el Instituto de Biotecnología de las Plantas de la Universidad Central de Marta Abreu de las Villas, Facultad de Ciencias Agropecuarias. Santa Clara Cuba. Wilder Camacho Chiu Doctor en Ciencias Agrícolas, Universidad Autónoma de Nuevo León, México. Maestro en Ciencias en Suelos Tropicales. Ingeniero Agrónomo, Colegio Superior de Agricultura Tropical, México. Correo electrónico: wildercamachochiu@hotmail.com Raúl Collado López Maestro por Instituto de Biotecnología de las Plantas. Universidad Central “Marta Abreu” de Las Villas. Felipe Jiménez Ferry Profesor Investigador por el Instituto de Biotecnología de las Plantas de la Universidad Central de Marta Abreu de las Villas, Facultad de Ciencias Agropecuarias. Santa Clara Cuba.
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
Marta Pérez Peralta Biotecnología Vegetal por el Instituto de Biotecnología de las Plantas de la Universidad Central de Marta Abreu de las Villas, Facultad de Ciencias Agropecuarias. Santa Clara Cuba. Odalys Gutiérrez Martínez Profesor Investigador por el Instituto de Biotecnología de las Plantas de la Universidad Central de Marta Abreu de las Villas, Facultad de Ciencias Agropecuarias. Santa Clara Cuba.
Forget SARS, West Nile, Ebola and avian flu. The real epidemic is fear. http://www.macleans.ca/shared/print.jsp?content=20051003_113117. Forget SARS, West Nile, Ebola and avian flu. The real epidemic is fear. We keep bracing ourselves for one cataclysmic threat after another. Our perceived lack of safety has become an obsession, Gregory Fields is a pharmaceutical maverick. He cal
Manuel Bernardo Barbosa - mbb@di.uminho.pt3. Specification and validation of CANopen using PROMELA and Spin4. Analysis of the Conformance Testing approachManuel Bernardo Barbosa - mbb@di.uminho.ptCANopen - Formal Specification and Validation - Conformance Testing• CANopen is a communication protocol designed for distributed controlin industrial environments: system integration. • Typica

 Ra Ximhai
Ra Ximhai  Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
EMBRIOGÉNESIS SOMÁTICA EN (Anthurium andraeanum Lind.) VARIEDAD
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
EMBRIOGÉNESIS SOMÁTICA EN (Anthurium andraeanum Lind.) VARIEDAD 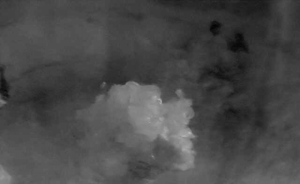 Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
prueba de rangos múltiples según Tukey y Dunnett’s con una significación de 0.05,
mediante el programa estadístico SPSS versión 13.0 para ‘Windows’.
RESULTADOS Y DISCUSIÓN
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
prueba de rangos múltiples según Tukey y Dunnett’s con una significación de 0.05,
mediante el programa estadístico SPSS versión 13.0 para ‘Windows’.
RESULTADOS Y DISCUSIÓN 
 Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
Figura 2. (A)- Callo con embriogénesis de alta frecuencia (ESAF) obtenido a partir de explantes foliares de
Ra Ximhai Vol. 4. Número 1, enero – abril 2008, pp. 135-149.
Figura 2. (A)- Callo con embriogénesis de alta frecuencia (ESAF) obtenido a partir de explantes foliares de