Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
170258.qxd
Int J Colorectal Dis (2000) 15:291–296DOI 10.1007/s003840000258
Bidirectional supply of glutamine maintains
J.D. Söderholm J. Larsson
enterocyte ATP content in the in vitro Ussing
J. Permert J. Lindgren M. Wirén Abstract Glutamine is the principal
Accepted: 19 September 2000Published online: 9 November 2000
(P<0.05), and the addition of gluta-
circuit current (P<0.05). No signifi-
H. Yang · J.D. Söderholm · J. Lindgren
Faculty of Health Science,Linköping University,
Keywords Ussing chamber ·
Karolinska Instituteat Huddinge University Hospital,
kept viable in Krebs solution containing glutamate interms of permeability and electrical properties, but the
One important feature of small-bowel function is to
effects of glutamine, the preferred energy source, have
maintain the barrier function while absorbing nutrients.
The function of tight junctions is vital to the barrier and
Glutamine is considered essential for the maintenance
can be measured by studying the permeation of markers
of gut metabolism, structure, and function in stress con-
that pass paracellularly. In vitro studies are valuable
ditions [4]. It plays an important role as a respiratory
tools for studying this phenomenon in specific parts of
substrate for cells in the mucose of the small intestine
the intestine under standardized conditions. In the Us-
[5,6]. Metabolism of this amino acid to α-ketoglutarate
sing chamber, transepithelial potential difference (PDt),
with subsequent complete oxidation in the Kreb’s cycle
short circuit current (Isc), and transepithelial electrical
yields 30 mol ATP per mol glutamine [7]. Enterocytes
resistance (TER) can be monitored over the membrane
are strongly dependent on an external glutamine supply
for optimal surveillance of the mucosa in different dis-
because of the small size of the cellular glutamine pool
ease states [1, 2, 3]. Stripped mucosal segments can be
in the small intestine [5]. In addition, the glutamine syn-
thetase activity in the intestinal mucosa is extremely low
both the mucosal and serosal side. The exposed tissue surface area
[8]. The mucosa is dependent on both luminal and plas-
was 1.78 cm2. The Krebs buffer was continuously oxygenatedwith O /CO (95/5%) and stirred by gas flow in the chambers.
ma glutamine and extracts 20–30% of circulating gluta-
After a 40-min equilibration period to achieve steady-state condi-
tions regarding PDt, the Krebs buffer in the serosal compartment
Glutamine is also an important nucleotide precursor
was replaced and marker solution containing 51Cr-EDTA with or
for the gastrointestinal tract [10, 11]. Using in vivopoly-
without glutamine was added to the mucosal compartments;
ethylene glycol (PEG) recovery as a measurement of epi-
0.6 mM glutamine corresponds to normal plasma levels of gluta-mine. The three groups (n=6 in each group) were treated as fol-
thelial absorptive and discriminatory function, we found
lows: KB group, Krebs buffer solution; 6 KB group, 6 mM gluta-
that glutamine supplementation resulted in an increased
mine on the mucosal side in the Krebs buffer solution; 0.6+6 KB
absorptive area of the small bowel [12]. Using the Us-
group), 6 mM glutamine on the mucosal side and 0.6 mM gluta-
sing chamber, we have found that starvation opens up
mine on the serosal side in Krebs buffer solution.
Samples of 1 ml were taken every 20 min from the serosal
the paracellular pathway for 51Cr-ethylenediaminetetra-
compartment for subsequent analysis of 51Cr-EDTA and were re-
acetate (EDTA). We have previously shown that starva-
placed by fresh Krebs buffer. Incubation was performed for
tion and surgery increase paracellular permeability and
180 min after equilibration, and specimens were then removed for
that starvation is the most influential factor in this exper-
imental model [13]. We have also examined the effectsof adding glutamine at different concentrations, i.e.,
0.6 mM, 3 mM, 6 mM, and 30 mM in Krebs solution
One pair of Ag/AgCl electrodes (Radiometer, Copenhagen,
without any nutrients, i.e. glutamate, glucose, fumarate,
Denmark) with 3 M KCl/2% agar bridges was used for measure-
and pyruvate, to the mucosal side of the jejunum of
ments of the transepithelial potential difference, and another pair
starved rats and found that this unidirectional supply of
of Pt electrodes was used for current passage. The electrodes werecoupled to an external six-channel electronic unit with a voltage-
glutamine increased ion pump activity but did not restore
controlled current source. Data sampling was computer controlled
the intracellular ATP level [14]. Whether an optimal glu-
via an A/D–D/A board (Lab NB, National Instruments) by a pro-
tamine plasma concentration or enteral administration of
gram developed in Lab View (National Instruments). A linear least
glutamine is needed to restore gut integrity in a metabol-
square fit was performed on the current (I) to voltage (U) pair re-
The purpose of this study was to determine the effects
of glutamine supplemented to one or both sides of the
The transepithelial resistance (TER) is obtained from the slope of
mucosa on the viability, energy metabolism, and epithe-
the line, potential difference (PDt) from the intersection of the
lial permeability of stripped jejunal mucosa from starved
voltage axis (when I=0), and the short circuit current (Isc) deter-mined from the quotient PD/TER. Three representative time
points, i.e., at equilibration point (0 min), 15 min (the effect ofadding glutamine to the system), and 180 min after equilibration(the long-term effect on cellular metabolism and viability) during
incubation in Ussing chambers were chosen for analysis.
Male Wistar rats (B&K Universal AB, Stockholm, Sweden),
Krebs buffer was prepared for each day of experiment and con-
weighing 240–260 g, were used. The rats were housed two by two
tained the following: NaCl 110.0 mM, CaCl 3.0 mM, KCl
in a room maintained at 23°C in a 12-h dark/light cycle throughout
all the experiments and were fed a normal diet with no restriction
5.7 mM, Na fumarate 7.0 mM, Na glutamate 5.7 mM, and glucose
on food or water supply for 2 weeks.
13.4 mM. It was adjusted to a pH of 7.3 and equilibrated with
Prior to the Ussing chamber experiments, the animals had no
access to food but free access to water for 48 h. Rats fed ordinary
Glutamine was prepared and dissolved in Krebs buffer before
chow who were not starved served as controls (n=6). Anesthesia
the Ussing chamber study. All the glutamine solutions were ad-
with ketamine (Ketalar, 80 mg/kg) and xylazine (Rompun,
justed to a pH of 7.3, and the addition of glutamine did not signifi-
8 mg/kg) was administered intraperitoneally. The abdomen was
opened by a full-length midline incision, and a 20-cm segment of
51Cr-EDTA (Du Pont, Dreieich, Germany), specific activity
the proximal jejunum was stripped from the mesentery and imme-
126 Ci/mmol, was added to the mucosal compartment at the start
diately put in ice-cold, oxygenated Krebs buffer. While immersed
of the experiments to a concentration of 44.3 µg/l (0.13 µM). Per-
in cold Krebs buffer, the outer muscle layers were removed under
meability of 51Cr-EDTA was assessed by measuring the appear-
a dissection microscope, and the tissue sections were cut into
ance of the marker on the serosal side during the experiments. The
specimens of an appropriate size and mounted in modified Ussing
radioactivity in 1-ml samples was counted for 600 s in a γ-counter
chambers [15) (Precision Instrument Design, Los Altos, CA,
(1282 Compugamma, LKB, Bromma, Sweden).
The apparent permeability coefficient (P
After mounting, each half cell was filled with 5 ml preheatedKrebs buffer at 37°C, bathing the stripped mucosa specimen on
(cm/s) = (dC/dt) × V(1/Co × A)
where dC/dt is the change in concentration on the serosal side perunit time (mol/l per s), V is the volume of the chamber (cm3), A isthe area of exposed intestine (cm2), and Co is the initial markerconcentration in the mucosal reservoir (mol/l) [16]. P
culated for the 20–120 min period.
Samples of jejunal mucosa were taken at operation as controlvalue (n=6) and after 180 min of incubation in the Ussing cham-ber. The samples were frozen in liquid nitrogen, stored at –70°C,and freeze dried. At analysis, mucosa powder was dissected freeof blood and connective tissue under a microscope and ground to ahomogenous powder. ATP was extracted from no less than 2.5 mgfreeze-dried mucosa powder using 1 M perchloric acid containing1 mM EDTA. The extracts were neutralized with 2.2 M KHCO3
and stored at –70°C until analysis. Analyses were performed with
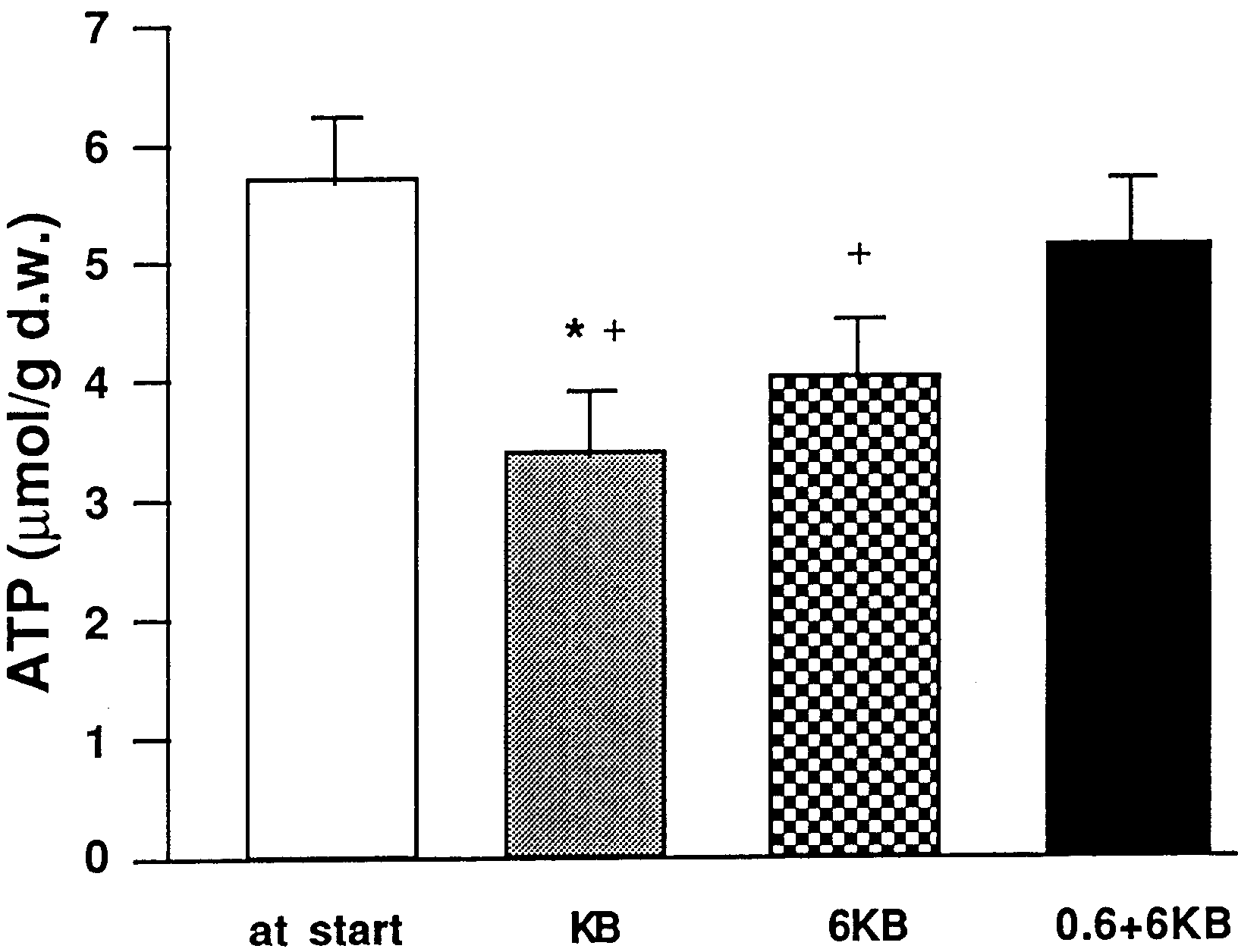
Fig. 1 Changes in ATP content of stripped jejunal mucosa during
enzymatic fluorometry using methods modified from Harris et al.
incubation in Ussing chambers. Values are expressed as mean ±
[17]. The contents of ATP were expressed as micromoles per gram
SEM micromoles per gram of dry weight mucosa tissue. *P<0.05
vs. 0.6+6 KB group, +P<0.05 6 KB vs. value at start
Data are presented as mean±standard error of mean (SEM). Com-parisons between groups were evaluated using the Kruskal Wallistest and in-group comparisons using the Wilcoxon’s paired signedrank test. Differences were considered significant at P<0.05.
The study was approved by the Animal Ethics Committee of theFaculty of Health Science, Linköping University, Sweden.
ATP contents of the stripped jejunal mucosa
When no glutamine was added (KB group), ATP de-
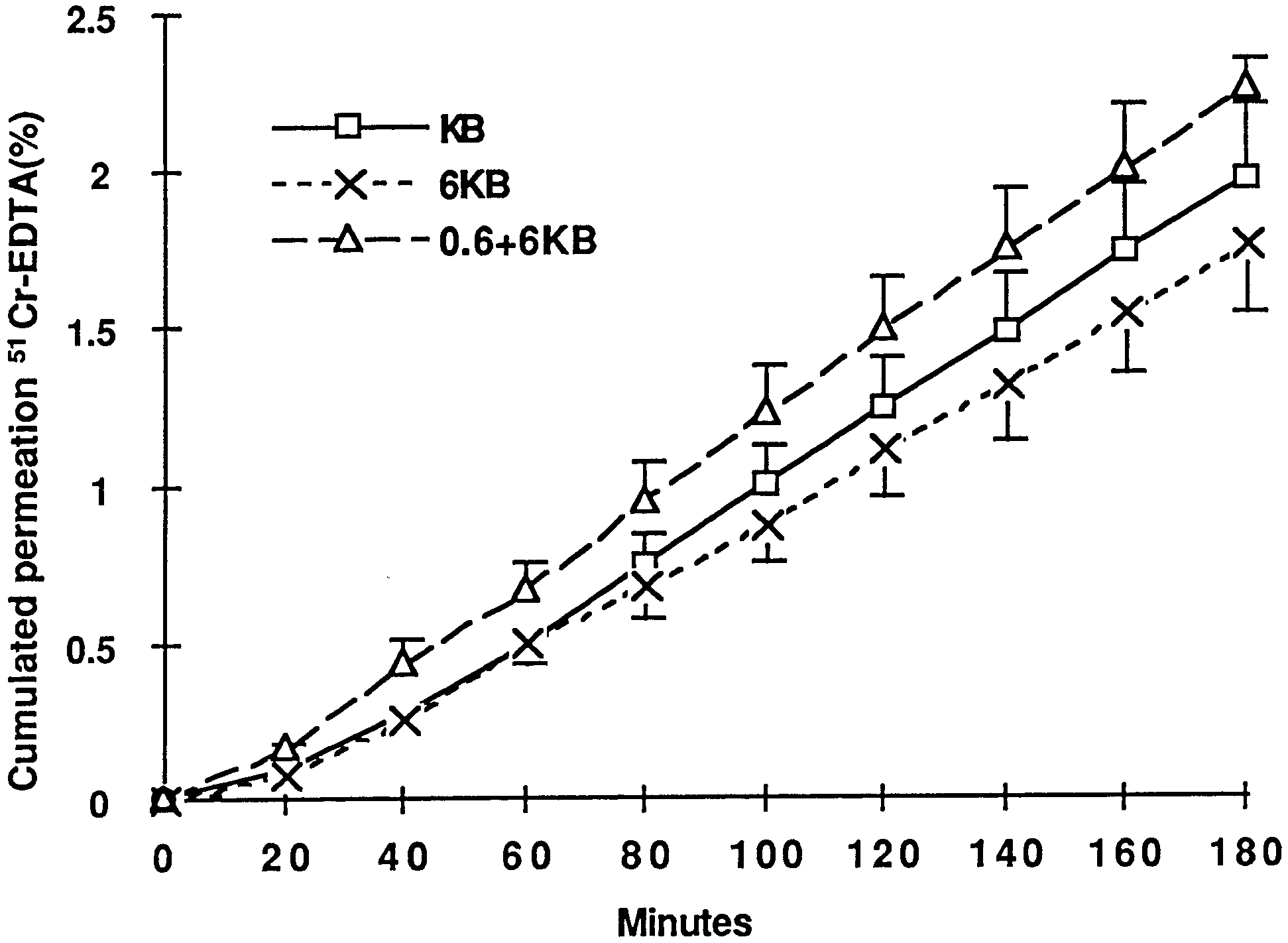
Fig. 2 Cumulative transmucosal permeation of 51Cr-EDTA in
creased by 40% compared to sampling at the start of ex-
stripped jejunal mucosa of starved rats specimens during 180 min
periment (3.4±0.5 vs. 5.7±0.5 µmol/g d.w., P<0.05). The
of incubation after equilibration in Ussing chambers. Vertical linesindicate SEM. There was a linear permeation increase during
mucosal ATP level when glutamine was added only on
180 min of incubation in all groups. Squares, KB; crosses, 6 KB;
the mucosal side was also significantly lowered
(4.0±0.5 µmol/g d.w., P<0.05). On addition of glutamineto both the mucosal and the serosal side, the ATP content(5.1±0.6 µmol/g d.w.) was maintained compared to the
The short circuit current (Isc) in stripped jejunal mucosaat equilibration was 131±12 µA/cm2 in the KB group vs.
138±22 µA/cm2 in the 6 KB group and 127±22 µA/cm2in the 0.6+6 KB group (not significant) (Figs. 3, 4). Rel-
The cumulated jejunal permeability of 51Cr-EDTA ative values during the experiment are presented inof starved rats did not differ significantly at 120 min Fig. 3. The addition of glutamine significantly increasedin the three groups (Fig. 1), and P
Isc after 15 min in the Ussing chamber(P<0.05), and bi-
in the 0.6+6 KB group, 5.5±0.7 cm/s in the KB group,
directional glutamine increased Isc more rapidly. Isc was
and 4.9±0.6 cm/s–1 in the 6 KB group (not significant).
also higher at 180 min in glutamine-containing KB
of intestinal mucosa found in previous experi-
ments in rats feeding normally is 3.3±0.3 cm/s [10]
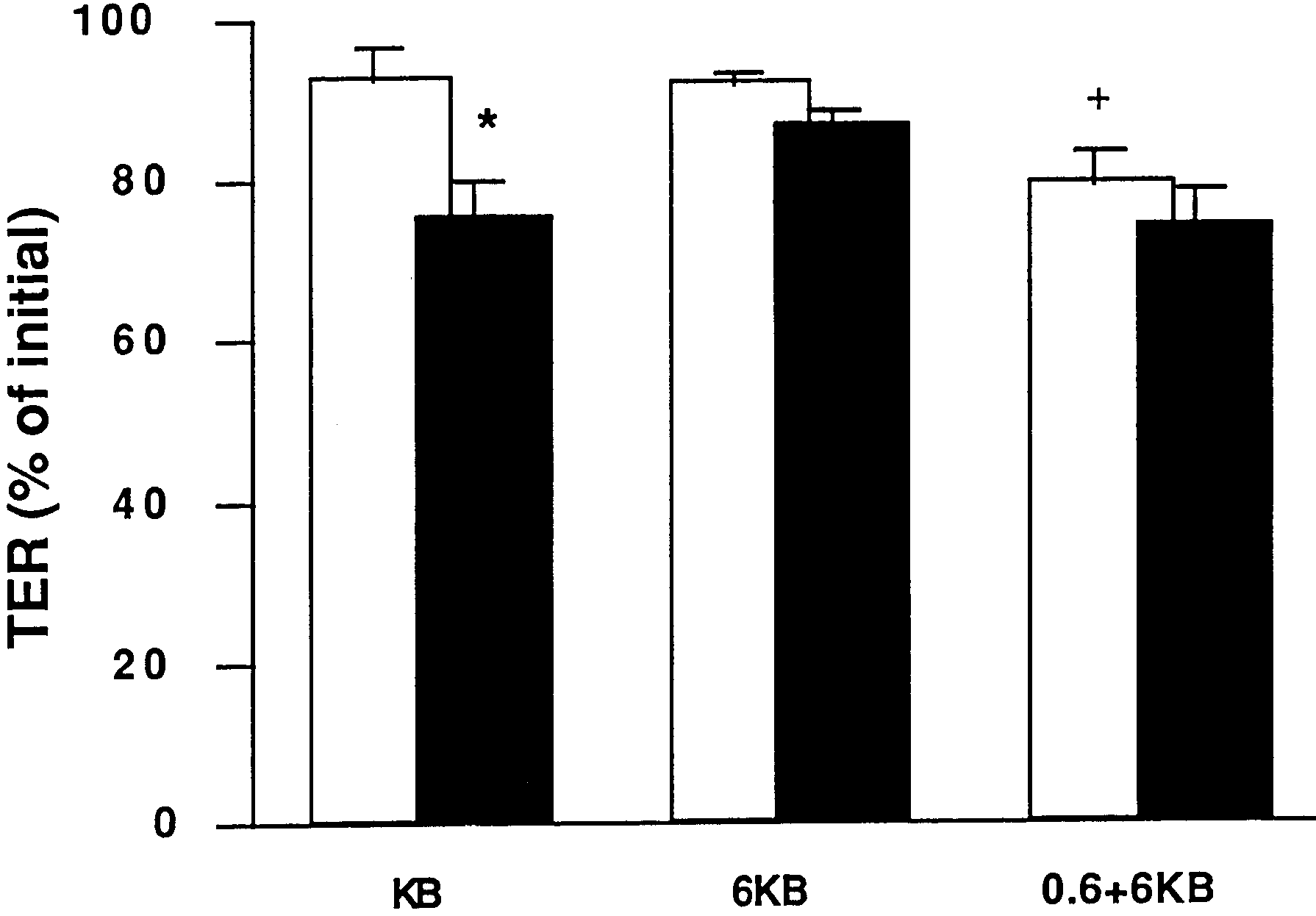
The TER in stripped jejunal mucosa at equilibration
was 26.2±3.2 Ωcm2in the KB group vs. 28.5±2.3 Ωcm2
interest. However, the optimal route of supply for gluta-mine is still under debate. To elucidate the specific roleof glutamine from the luminal side or both the luminaland serosal side, we used the Ussing chamber model tostudy the effects on the energy metabolism and perme-ability of stripped intestinal mucosa. We found that theaddition of glutamine supports ATP levels and ion pumpactivity to a higher degree when glutamine is providedfrom both sides of the mucosal lining. We suggest thatthe supply of a physiologic concentration of glutamine tothe serosal side is important for maintenance of mucosalviability.
Our hypothesis was that glutamine is beneficial to en-
terocyte energy metabolism in the stressed situation inthe Ussing chamber. We have previously reported bene-
Fig. 3 Changes in short-circuit current (Isc) in jejunal mucosa of
ficial effects of glutamine on proliferation, but these
starved rats in a Ussing chamber. White bars, 15 min after equili-
effects are only demonstrated when there is depletion or
bration; black bars, 180 min after equilibration. Data are ex-
an increased demand of glutamine compared to the fed
pressed as mean±SEM percentage of the initial value at equilibra-tion time. *P<0.05 vs. KB group at 15 min; +P<0.05 vs. KB group
state. For this reason, starved animals were studied. Glu-
tamine has been suggested to be particularly efficientwhen glycolysis is depressed [18]. It has recently beenshown that glutamine supplement can increase transmu-cosal resistance and decrease mannitol flux through theepithelium in a severe acute colitis model in an Ussingchamber [19]. Keurkchubascke et al. [20] found that in-testinal membrane perfused with a solution consisting ofDulbecco’s modified Eagle’s medium with 20 mM gluta-mine maintained the TER for 3 h in an Ussing chamber. In our study, we found that the permeability perturbationresulting from starvation was not attenuated by addingglutamine to the Ussing chamber. This might be due tostructural changes rather than to an energy deficit in theepithelial lining. TER is thought to reflect tissue integrity. A low TER value suggests increased permeability to ionicmovement or lower tissue integrity. When glutamine wasadded to both sides, there was a rapid drop in TER,
Fig. 4 Changes in transepithelial resistance (TER) in jejunal mu-
which might be caused by an increase in ionic flux due
cosa of starved rats in vitro. White bars, 15 min after equilibration;
to activation of Na+-coupled absorption of glutamine,
black bars, 180 min after equilibration. Data are expressed asmean±SEM percentage of the initial value at equilibration time
since Isc also increased more rapidly in this group at this
point. *P<0.05 15 min vs. 180 min within group; +P<0.05 vs. KB
time. In this short experiment, tight junction permeation
might have been initially affected by glutamine, butthere were no significant differences in TER or P
in the 6 KB group and 27.6±2.8 Ωcm2 in the 0.6+6 KB
the end of the experiment among the three groups.
group (not significant). There was a significant drop in
Adenine nucleotide levels indicate viability of intes-
TER after 15 min in the 0.6+6 KB group and in the KB
tine [21], and ATP levels in enterocytes appears to be
group (Fig. 4). There were no significant differences in
important for the maintenance of intestinal barrier func-
TER between the three groups at the end of the experi-
tion [22]. The importance of ATP for maintaining normal
permeability characteristics in epithelial monolayers [23,24, 25] has been extensively documented. Profound ATPdepletion led to loss of both the “gate” and “fence” func-
tions of tight junctions. In both endothelial cells and kid-ney epithelia, ATP depletion results in a redistribution of
Starvation and surgical stress increase the vulnerability
cortical membrane-associated filamentous actin [26, 27,
of the intestinal mucosa and might interfere with barrier
28, 29]. In the study presented here, glutamine on both
integrity. Specific nutrients to support mucosal energy
the luminal and serosal side demonstrated the ability to
restoration and to stimulate proliferation are of clinical
preserve the energy charge, and ATP content in stripped
mucosa only decreased by 10% of the initial value when
In summary, this study suggests that supply of gluta-
glutamine was added to both sides. In contrast, the gluta-
mine from both the luminal and serosal side seems to be
mate and glucose content of the Krebs buffer was unable
important for the maintenance of energy metabolism and
to maintain adenine nucleotide levels, and ATP content
mucosal viability in rat intestinal mucosa. The two meth-
decreased by 40% of the initial value when glutamine
ods of administration might result in different processing
of the supplemented amino acid with regard to transport
The ability to maintain an Isc is a characteristic
mechanisms versus oxidation. The possibility of there
shared by all transporting epithelia and is dependent on
being a differentiated metabolic cellular response to par-
the electrogenic ion pumps in the epithelial function
enteral and enteral glutamine needs further attention.
[30]. Thus Isc is equivalent to the sum of all active ion
The addition of glutamine to the mucosal side and a
transport processes which require energy production,
physiologic plasma concentration of glutamine to the
generally in the form of ATP. Tissue viability in small-
serosal side might also be a valuable model to improve
intestine tissue can also be evaluated from the basal Isc
intestinal energy metabolism in future in vitro studies.
or from the changes in Isc when resistance is stable [31]. Acknowledgement This study was supported in part by the Med-
A low Isc suggests that the stripped mucosa has a low
ical Research Council (10402 and 12618) and grants from the
metabolic rate or ionic flux. In our study, Isc was higher
County Council of Östergötland and Semper AB. We are indebted
in all groups when the Krebs buffer was supplemented
to Ms. B. Ylva for the analysis of the energy metabolism. Dr. Hua
with glutamine, especially bidirectionally. Adding gluta-
Yang is a Research fellow from the Department of Surgery,Xinqiao Hospital, Chongqing 630037, China. He is currently work-
mine might therefore be a way to increase cellular viabil-
ing at the Section of Pediatric Surgery, University of Michigan
Hospitals, Mott F3970, Box 0245, Ann Arbor, MI 48109, USA.
Altered tight junction structure contrib-
function in ulcerative colitis. Gastroen-
tional factors and bacterial virulence.
21. Hirata Y, Taguchi T, Suita S, Takeshige
lism in relation to graft viability in the
vivo for post absorptive rat small intes-
and permeability of goblet cell tightjunctions in rat small intestine. J Membr Biol 66:145–157
‘fence’ and paracellular ‘gate’ func-
tions in epithelial tight junctions.
logic impact of cellular oxidant injury.
membrane actin cytoskeletal complexduring cellular ATP depletion. J ClinInvest 88:462–469
Microfilament disruption occurs veryearly in ischemic proximal tubule in-jury. Kidney Int 42:896–902
My pregnancy went without a hitch. I had no morning sickness or nausea or any other symptoms to trouble me. As a result I was completely calm and relaxed throughout. I went overdue, and at 41+6 was taken into hospital to be induced. I was hooked up to a monitor that indicated I was having contractions, although I couldn’t feel them. I was sent home shortly after being monitored as Labour Ward w
 where dC/dt is the change in concentration on the serosal side perunit time (mol/l per s), V is the volume of the chamber (cm3), A isthe area of exposed intestine (cm2), and Co is the initial markerconcentration in the mucosal reservoir (mol/l) [16]. P
culated for the 20–120 min period.
where dC/dt is the change in concentration on the serosal side perunit time (mol/l per s), V is the volume of the chamber (cm3), A isthe area of exposed intestine (cm2), and Co is the initial markerconcentration in the mucosal reservoir (mol/l) [16]. P
culated for the 20–120 min period.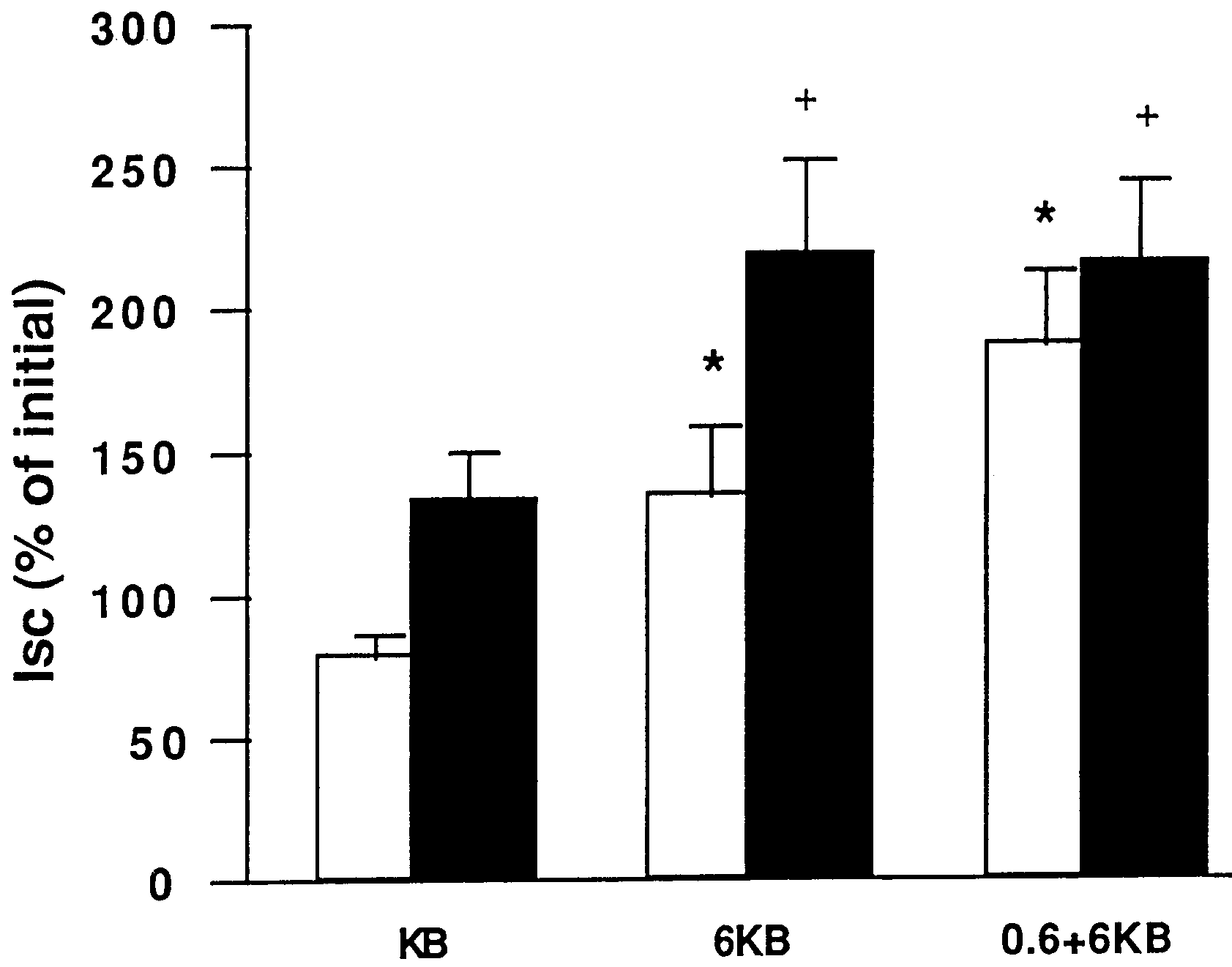
 interest. However, the optimal route of supply for gluta-mine is still under debate. To elucidate the specific roleof glutamine from the luminal side or both the luminaland serosal side, we used the Ussing chamber model tostudy the effects on the energy metabolism and perme-ability of stripped intestinal mucosa. We found that theaddition of glutamine supports ATP levels and ion pumpactivity to a higher degree when glutamine is providedfrom both sides of the mucosal lining. We suggest thatthe supply of a physiologic concentration of glutamine tothe serosal side is important for maintenance of mucosalviability.
interest. However, the optimal route of supply for gluta-mine is still under debate. To elucidate the specific roleof glutamine from the luminal side or both the luminaland serosal side, we used the Ussing chamber model tostudy the effects on the energy metabolism and perme-ability of stripped intestinal mucosa. We found that theaddition of glutamine supports ATP levels and ion pumpactivity to a higher degree when glutamine is providedfrom both sides of the mucosal lining. We suggest thatthe supply of a physiologic concentration of glutamine tothe serosal side is important for maintenance of mucosalviability.