Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Rborges.webs.ull.es
0022-3565/02/3012-631–637$7.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS
Copyright 2002 by The American Society for Pharmacology and Experimental Therapeutics
Nongenomic Regulation of the Kinetics of Exocytosisby Estrogens
JOS ´E D. MACHADO, CARMEN ALONSO, ARACELI MORALES, JOS ´E F. G ´
Unidad de Farmacologı´a (J.D.M., C.A., J.F.G., R.B.) and Laboratorio de Neurobiologı´a Celular (A.M.), Facultad de Medicina, Universidad de LaLaguna, Tenerife, Spain
Received November 2, 2001; accepted February 1, 2002
This article is available online at http://jpet.aspetjournals.org
ABSTRACT The role of nongenomic action of estrogens on elicited cat-
down of the exocytosis as measured by amperometry.
echolamine secretion and exocytosis kinetics was studied in
perfused rat adrenals and in cultured bovine chromaffin
echolamines. 17-Estradiol reduced the intracellular free
cells. 17-Estradiol as well as the estrogen receptor modu-
Ca2ϩ but only at micromolar concentrations, whereas nano-
lators raloxifene and LY117018, but not 17␣-estradiol, inhib-
molar concentrations increased the cAMP levels. These ef-
ited at the micromolar range the catecholamine output elic-
fects were reproduced with the nonpermeable drug 17-
ited by acetylcholine or high potassium. However, these
agents failed to modify the secretion elicited by high Ca2ϩ in
nanomolar concentrations of the antiestrogen ICI 182,780
glands treated with the ionophore A-23187 (calcimycin), sug-
(fulvestrant). Our data suggest the presence of membrane
gesting that estrogens did not directly act on the secretory
sites that regulate both the exocytotic phenomenon and the
machinery. At the single cell level, estrogens modified the
total catecholamine release with high and low affinity, re-
kinetics of exocytosis at nanomolar range. All of the drugs
tested except 17␣-estradiol produced a profound slowing
The nongenomic actions of estrogens are receiving a re-
observation has motivated the coining of the term “estrogen
newed interest. It is now becoming clear that the rapid onset
modulator” for tamoxifen and other compounds like ralox-
of cellular responses upon drug application, together with the
ifene or LY117018, which are currently under investigation
insensitivity of these responses to blockers of transcription or
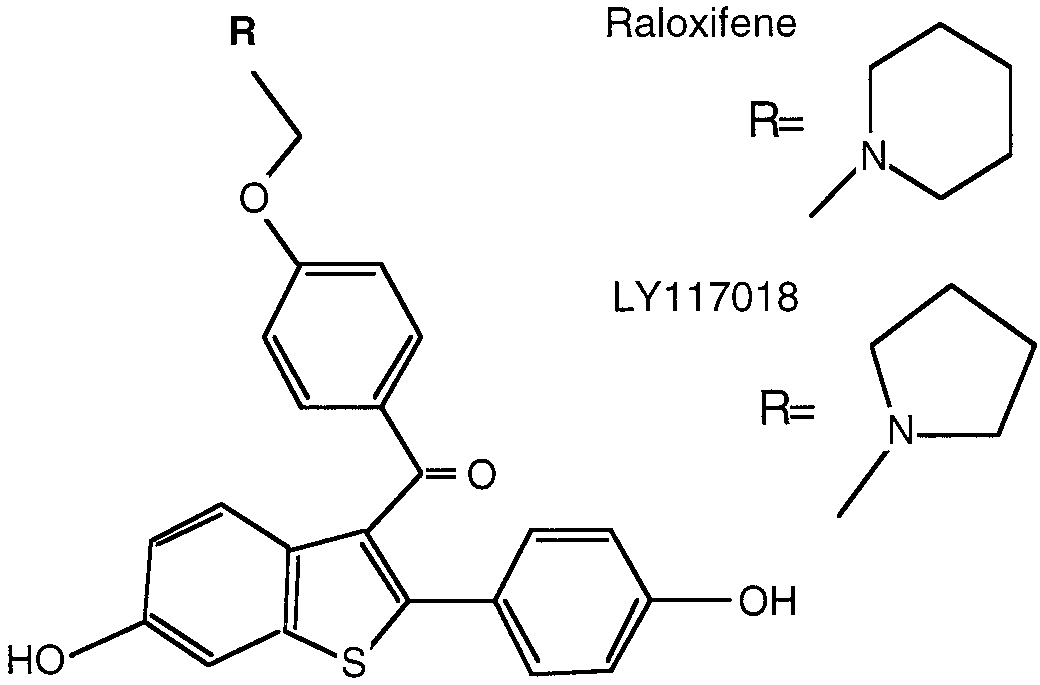
(Fig. 1). The reasons explaining their different tissue selec-
translating, cannot be attributed to genomic activity (Weh-
tivity and specific activity are still obscure and cannot be
satisfactorily explained by the simple ␣- and -estrogen re-
The naturally occurring steroid 17-estradiol, its ␣-isomer,
ceptor affinity (Nadal et al., 2000).
and other molecules with estrogen activity have been widely
The sites of action responsible for the rapid action of ste-
studied in a variety of tissues including chromaffin cells
roids are a source of controversy. In an attempt to provide an
(Lo´pez et al., 1991; Park et al., 1996; Dar and Zinder, 1997)
assessment of the nongenomic receptors, the so-called Mann-
and PC-12 cells (Chen et al., 1998; Kim et al., 2000; for a
heim classification was published (Falkenstein et al., 2000a);
recent review, see Falkenstein et al., 2000b).
however, it says very little about the nature of receptors
A number of drugs with estrogenic activity have been syn-
involved in the rapid cellular responses to steroids. One of
thesized; some of them exhibited agonist effects in some
the candidates for being a receptor for estradiol is the -sub-
tissues whereas they behaved as antagonists in others. This
unit of the maxi-K potassium channel (Valverde et al., 1999). However, the presence of functional classical nuclear recep-tors on cell membrane has been described (Watson et al.,
This work was supported in part by grants from Spanish Ministerio de
1999; Razandi et al., 2000; Wyckoff et al., 2001), although
Ciencia y Tecnologı´a (PB97-1483 and BFI2001-3531), Gobierno de Canarias,and Fondo Europeo de Desarrollo Regional (1FD97-1065-C03-01). We also
they are far from explaining all of the membrane-mediated
received partial financial support from Eli Lilly & Co. SA (Madrid), Zeneca
effects thus far described for estrogens.
Farma, SA (Madrid), and Compania Espanola de Petro´leos Sociedad Anonima(Tenerife). J.D.M. was the recipient of a fellowship from Instituto Tecnolo´gico
Several second messenger routes were implicated in the
de Canarias, A.M. from Spanish Ministerio de Ciencia y Tecnologı´a, and J.F.G.
cellular signaling occurring upon nongenomic estrogen stim-
from Consejerı´a de Educacio´n del Gobierno de Canarias. A poster from this
ulation. These routes included cAMP (Minami et al., 1990;
paper was presented at the 11th International Symposium on Chromaffin CellBiology (San Diego, CA).
Gu and Moss, 1996), inositol 1,4,5-trisphosphate formation
ABBREVIATIONS: ACh, acetylcholine; CA, catecholamine; [Ca2ϩ] , cytosolic calcium concentration; DMPP, 1,1-dimethyl-4-phenylpiperazinium;
HRP, horseradish peroxidase; IBMX, 3-isobutyl-1-methylxanthine; ICI 182,780, fulvestrant; A-23187, calcimycin. Machado et al.
Guide for the Care and Use of Laboratory Animals and approved byThe Ethical Committee of La Laguna University (Tenerife, Spain).
To establish the possible role of estrogens on the secretory ma-
chinery, we performed some experiments in A-23187 permeabilizedtissues. Glands were perfused with Krebs’ solution lacking Ca2ϩ(CaCl was isosmolarly replaced by MgCl ) for 15 min, then a 10-M
solution of A-23187 was perfused for 10 min. Glands were perfused inCa2ϩ-lacking solution, and the secretion was triggered by risenCaCl in the perfusate to 5 mM in 1-min duration pulses.
Ten minutes before and during the second stimulus, the glands
received 1 M estrogen; this concentration was raised to 10 M inthe third stimulus. Comparisons were made with the second andthird secretory response in the absence of drug. Statistical analysiswas performed with Dunnett’s paired t test. Culture Chromaffin Cells. Bovine adrenal chromaffin cells
were isolated as described previously (Moro et al., 1990) and platedon glass coverslips 12-mm in diameter at an approximate density of
Fig. 1. Structures of raloxifene and LY117018.
5 ϫ 104 cells/coverslip. Cells were used at room temperature between1 and 4 days of culture.
(Favit et al., 1991; Shears, 1991), and Ca2ϩ-current inhibi-
Amperometric Detection of Exocytosis. Carbon fibers of 5-m
tion (Ruehlmann et al., 1998; Kim et al., 2000). Recently,
radius (Thornel P-55; Amoco Corp., Greenville SC) were used to
Wyckoff et al. (2001) demonstrated the presence of a coupling
make the microelectrodes (Kawagoe et al., 1993). Electrochemical
between the ␣-receptor subtype and a nitric oxide synthase,
recordings were performed using an Axopatch 200B (Axon Instru-
but there are no studies available for other second messenger
ments, Union City, CA) (see Machado et al., 2000 for details).
Glass coverslips with adhering adrenal cells were washed in
cascades and their relationship with secretion.
Krebs-HEPES buffer solution containing 140 mM NaCl, 5 mM KCl,
It is important to make a distinction between effects on
1.2 mM MgCl , 2 mM CaCl , 11 mM glucose, and 10 mM HEPES,
secretion, i.e., the total amount of neurotransmitter release
brought to pH 7.35 with NaOH. Cells were placed in a perfusion
after a stimulus, and effects on exocytosis, i.e., kinetics at
chamber positioned on the stage of an inverted microscope. Ampero-
single event level. Both are closely related processes involved
metric measurements were performed with the carbon fiber micro-
in the release of neurotransmitters and other substances, but
electrode gently touching the cell membrane. Catecholamine release
the exocytotic kinetics can be drastically changed even when
was stimulated by 5-s pressure ejection of 5 mM Ba2ϩ, 10 M DMPP,
the total amount of products secreted results unchanged
or 59 mM Kϩ from a micropipette placed 40 m away from the cell.
(Machado et al., 2000). In addition, exocytosis can also occur
Amperometry Data Analysis. Amperometric records were low-
without secretion (Borges et al., 1997; Tabares et al., 2001).
pass filtered at 1 KHz, sampled at 4 KHz, and collected using a
In this study, we have addressed the question of whether
locally written software using LabVIEW for Macintosh (National
the role of estrogens in the regulation of secretory responses
Instruments, Austin, TX). To analyze the exocytotic events, a series
is related to changes in the kinetics of exocytosis or granule
of kinetics parameters were extracted from each spike. Data analysiswas carried out using locally written macros for IGOR (Wavemetrics,
content of catecholamine (CA). We have shown that estro-
Lake Oswego, OR). These macros allowed the automatic digital
gens, through a nongenomic mechanism, can modulate both
filtering, secretory spike identification, and data analysis (Segura et
processes, but with very different sensitivities. To our knowl-
al., 2000). All the above macros and their user instructions can be
edge, this is the first report describing the effects of estrogens
downloaded free from the following web address: http://webpages.
at the level of the single event of exocytosis.
In this study, significant differences were observed between the
untreated cells, used as controls, from different days and electrodes. Experimental Procedures Materials. Urografin was obtained from Schering Espan
to 88.1 pA. For this reason, effects of drugs on secretory spikes were
drid, Spain). Culture plates were purchased from Corning (Palo Alto,
always compared with control experiments carried out along the
CA). LY117018 and LY139481 (raloxifene) were a gift of Eli Lilly &
same day and using the same electrode. To avoid misinterpretation
Co. SA (Madrid, Spain). ICI 182,780 was a gift of Zeneca Farma, SA
of data, amperometric spike characteristics were not pooled but
(Madrid, Spain). All other drugs, culture media, and sera were pur-
grouped by individual cells (Colliver et al., 2000). Statistical analysis
chased from Sigma-Aldrich (Madrid, Spain). All salts used for buffer
was carried out by Mann-Whitney test. Measurement of Cytosolic-Free Ca2؉ Concentrations. Glass Perfused Rat Adrenals. Male Sprague-Dawley rats, weighing
coverslips with adhering cells were washed twice in Krebs’ buffer
200 to 300 g, were anesthetized with 50 mg/kg sodium pentobarbi-
solution and incubated with 2 M fura-2/acetoxymethyl ester (stock
tone i.p. Adrenal glands were perfused retrogradely in vitro at 1
solution dissolved in 20% pluronic F-127 gel in dimethyl sulfoxide)
ml/min, as described previously (Borges, 1993), with a Krebs-bicar-
and 0.1% fetal calf serum for 60 min at room temperature. Then, cells
bonate solution containing 119 mM NaCl, 4.7 mM KCl, 1.2 mM
were washed twice to remove extracellular dye and placed in a
MgSO , 1.2 mM KH PO , 2.5 mM CaCl , 25 mM NaHCO , and 11
perfusion chamber. Intracellular Ca2ϩ was measured using a com-
mM glucose. The pH was kept at 7.4 by continuous bubbling with
puter-operated monochromator (TILL Photonics, Munich, Germany)
95% O and 5% CO . The CA release was measured fluorimetrically
controlled by a locally written software using LabVIEW. Fluores-
by the trihydroxyindol method without further alumina purification
cence signals were low-pass filtered at 510 nm and detected by a
(Anton and Sayre, 1962). Glands received three consecutive stimuli
with ACh (30 M) or Kϩ (35.4 mM) of 1-min duration separated by 30
Data of [Ca2ϩ] time courses were collected at 1 Hz and expressed
min. All experiments were done at 37°C. All animal procedures were
made in strict accordance with the National Institutes of Health
ried out by Student’s t test. Estrogens and Exocytosis of Catecholamines cAMP Measurements. Cells were cultured on 24-well plates at
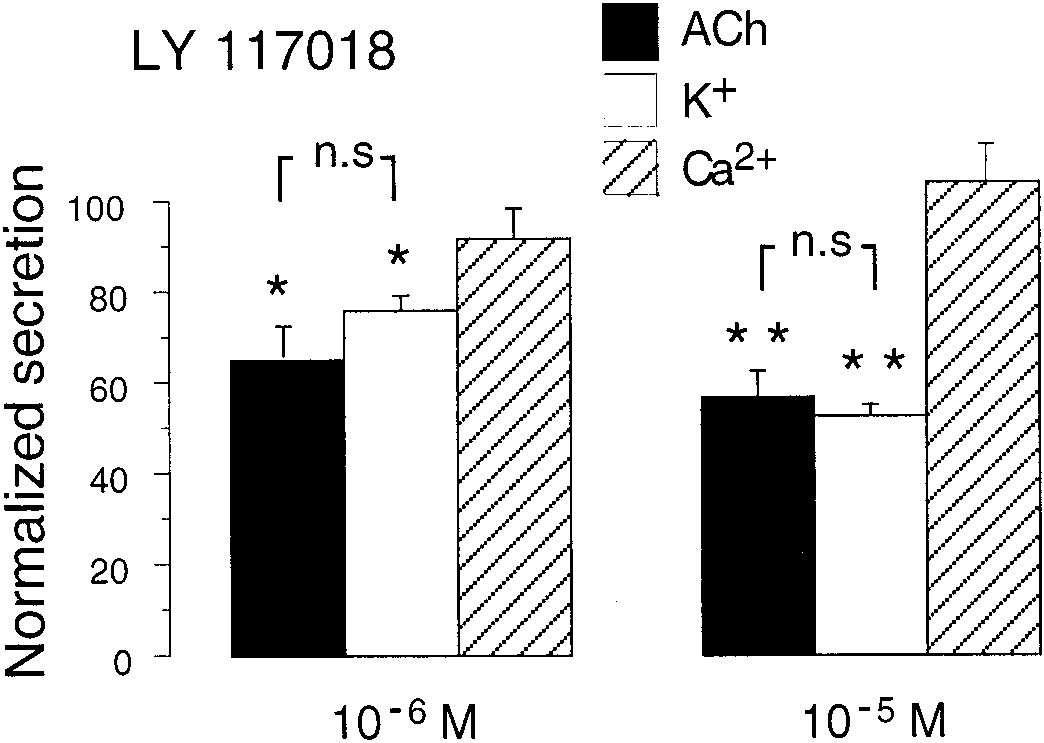
5 ϫ 105/well for 48 h. Cells were preincubated in Krebs-HEPESbuffer containing 500 M 3-isobutyl-1-methylxanthine (IBMX) for 15min. Testing drugs were incubated for another 15 min, always in thepresence of IBMX. Cyclic AMP measurements were done with thecAMP enzyme immunoassay (RPN225) kit (Amersham Biosciences,Cerdanyola, Spain). Data were expressed in femtomoles per micro-gram of total protein measured by the bicinchoninic acid method. Statistical analysis was carried out by a two-way analysis of variancefollowed by Tukey’s test. Estrogens Inhibited Secretion from Perfused Rat Adrenals in the Micromolar Range. Adrenal secretory responses to all of the secretagogues used were stable and reproducible along control experiments. Control experiments consisted of three repetitive stimuli with ACh (30 M; n ϭ 8), Fig. 3. Comparison of the effects of LY117018 on CA release evoked by
high Kϩ (35.4 mM; n ϭ 10), or high Ca2ϩ (5 mM, in glands
various agents on rat adrenals. Experiments were performed as described
pretreated with 10 M Ca2ϩ ionophore A-23187; n ϭ 7).
under Materials and Methods. Data (mean Ϯ S.E.M.) were compared
A series of estrogen molecules, 17␣-estradiol, 17-estradiol,
with the second or third stimuli (normalized as 100%) obtained in theabsence of drug. ૺ, p Ͻ 0.05; ૺૺ, p Ͻ 0.01 (Dunnett’s paired t test). Data
raloxifene, and LY117018, were assayed on ACh-evoked re-
were from 7 to 10 different glands of each group. No significant (n.s.)
sponses. None of these agents inhibited secretion at concentra-
differences were found between the inhibition observed on ACh and Kϩ
tions of 10Ϫ7 M or lower (data not shown). Inhibition of secre-
tion became evident over 10Ϫ6 M, although it was onlysignificant for LY117018. Raloxifene and 17-estradiol signifi-
only observed at 1 M (12% inhibition) and 10 M (38%
cantly attenuated the responses when given at 10 M; however,
17␣-estradiol failed to reduce the CA output (Fig. 2). No differ-
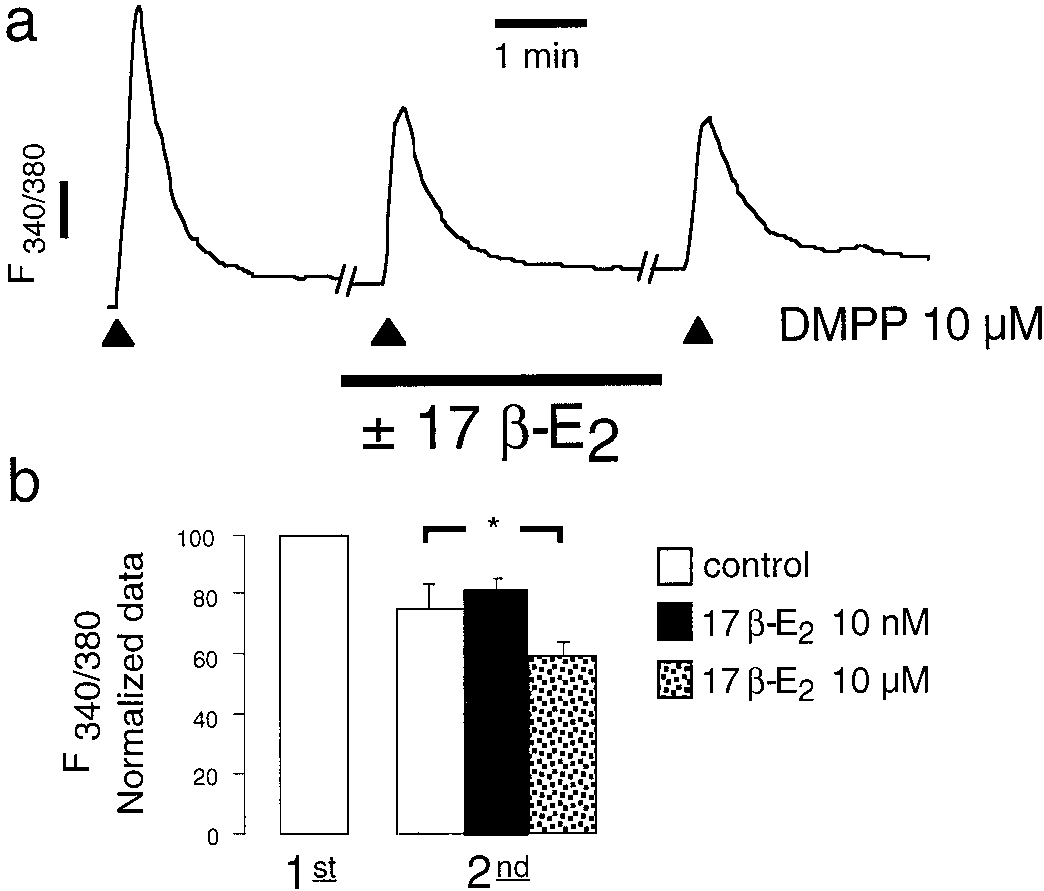
17-Estradiol Reduced the Ca2؉ Entry in the Micro-
ences in the degree of the blockade of secretion were observed
molar Range. Figure 4a shows a typical trace of the effect of
when 10 M was perfused without a previous incubation with
DMPP (10 M), applied for 5 s, on [Ca2ϩ] in an isolated bovine
lower concentrations of drugs (data not shown).
chromaffin cell. The nicotinic agonist applications were re-
Subsequent experiments were carried out to elucidate the
peated three times at 3-min intervals. Under control conditions,
possible cellular targets for estrogens. The LY117018 inhib-
desensitization occurred and the response fell down to 78 and
ited the secretion elicited with ACh or Kϩ in a similar extent
54%, respectively, on the second and third stimuli. Figure 4b
whereas it failed to modify the Ca2ϩ-evoked responses (Fig.
showed the average responses from six different control cells or
3). These data suggest that estrogen did not directly act on
after 10-min incubation with 17-estradiol at 10 nM or 10 M.
the secretory machinery but through a step situated betweencell membrane depolarization and the activation by Ca2ϩ ofthe secretory machinery.
The compound LY117018 also reduced the secretion of CA
from isolated bovine chromaffin cells when they were stimu-lated with 5-s stimuli separated by 5 min with high Kϩ (35mM) or DMPP (10 M). As in rat adrenals, this effect was
Fig. 4. The effects of low and high concentrations of 17-estradiol on Fig. 2. The effects of estrogens on acetylcholine-evoked responses. Estro-
[Ca2ϩ] . Bovine chromaffin cells were loaded with fura-2, as described
gens were applied along the second and third ACh stimuli at the concen-
under Materials and Methods, and stimulated with 5-s pulses of DMPP
trations shown. Data (mean Ϯ S.E.M.) were compared with the second or
(filled triangles) every 3 min. The estrogen was incubated from 10 min
third stimuli (normalized as 100%) obtained in the absence of drug. Data
prior to the second pulse and remained bathing cells along the rest of the
were pooled from 6 to 10 different glands of each group. ૺ, p Ͻ 0.05; ૺૺ,
experiment. a, typical fluorescent trace of 340:380 nm ratio; b, means Ϯ
p Ͻ 0.01 (Dunnett’s paired t test).
S.E.M. of six different cells from each group. ૺ, p Ͻ 0.05 (Student’s t test). Machado et al.
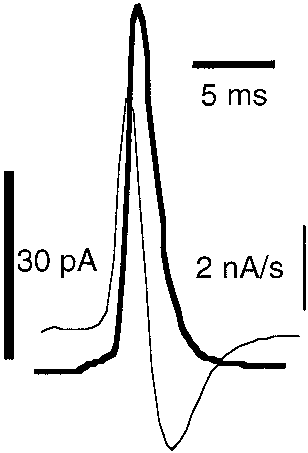
TABLE 1The effects of 17-estradiol and raloxifene on secretory spike kineticsInset figure describes the kinetic parameters measured. Imax is the maximal current caused by the CA reaching the electrode; t1/2 is the spike width at its half height; Q isthe integrated area under the spike trace that indicates the total CA released during the exocytotic event; m is the ascending slope, horizontal ticks indicate the 25 and 75%of Imax where m is calculated; tP is the time need to reach the spike maximum. Secretory spikes from cells treated 10 min with 10 nM estrogens are compared with untreatedcontrol groups. Data are expressed in the units indicated. Data were calculated by averaging spike parameters from each cell (means Ϯ S.E.M.). The number of spikescomputed was not taken into account for statistic analysis.
* Statistic differences (p Ͻ 0.05) from controls (Mann-Whitney tests).
This inhibitory effect of estrogen was only evident at micromo-
the presence of a foot (prespike feature) or the deceleration
lar concentrations. Similar effects were also observed with
observed just before the tip of spikes. Note that the maximum
raloxifene and LY117018 (data not shown).
releasing speed, in normal spikes, occurred at 52 Ϯ 0.5% of
Estrogens Affected the Kinetics of Exocytosis at the
, whereas the maximum acceleration took place at
Single Granule Level. It was only necessary for the brief
15 Ϯ 0.6%. Estrogens drastically reduced both the initial
application of nanomolar concentrations of estrogens to pro-
duce the slowing down of exocytosis. Table 1 shows the effects
Cell incubation with low concentrations of raloxifene (10
of 10 nM 17-estradiol or raloxifene on the kinetic parame-
nM) did not significantly modify the frequency of secretory
ters of single exocytotic events. Since the electrode was
spikes (90 Ϯ 13, n ϭ 10 versus 76 Ϯ 4, n ϭ 12 spikes).
placed onto the cell membrane, the slowing down of exocyto-
However, larger concentrations (1 and 10 M) caused also a
sis drug was causing an average reduction of a 38% of the CA
reduction in the number of exocytotic events. This reduction
concentration reaching its surface. Several estrogens repro-
was similar to that observed in net CA released from per-
duced this effect (Fig. 5), which seems to be mediated by a
membrane-associated receptor(s) since the cell-impermeable
The effects of raloxifene on the kinetics of exocytosis were
HRP-conjugated 17-estradiol caused similar effects. It is
studied at three different concentrations. Surprisingly, the
important to stress that 17-estradiol altered the kineticparameters, t
, m, and tP, but did not produce signif-
icant changes in net granule content of CA. All of these
The effects of various drugs with estrogen activity on ascending slope(m) of spikes
effects were observed within seconds of incubation.
Data (means Ϯ S.E.M.) are expressed in nanoamperes per second obtained in the
In Table 2, we analyzed how these compounds affect the
absence and presence of estrogens. % indicates the fall caused by drugs. Figure inset
initial speed of granule emptying. The figure accompanying
represents a spike with its first derivative (thinner trace) superimposed. Note thedifferent scale.
the table shows a spike with its first derivative trace. Theascending slope (m) was estimated between the 25 and 75%of the I
(Table 1, figure inset) to avoid interference from
Fig. 5. The effects of various estrogens on I
(means Ϯ S.E.M.) were compared with their own control group of
untreated cells (n ϭ 12–16). ૺ, p Ͻ 0.05 (Mann-Whitney test).
* p Ͻ 0.05 with respect to their own control (Mann-Whitney test). Estrogens and Exocytosis of Catecholamines
it was applied alone or in the presence of 17-estradiol. Thiscompound at 10 nM produced the slowing down of the exo-cytotic process, which was not accompanied by changes in theapparent granule content of CA. Estrogens Increased the Intracellular cAMP Levels.
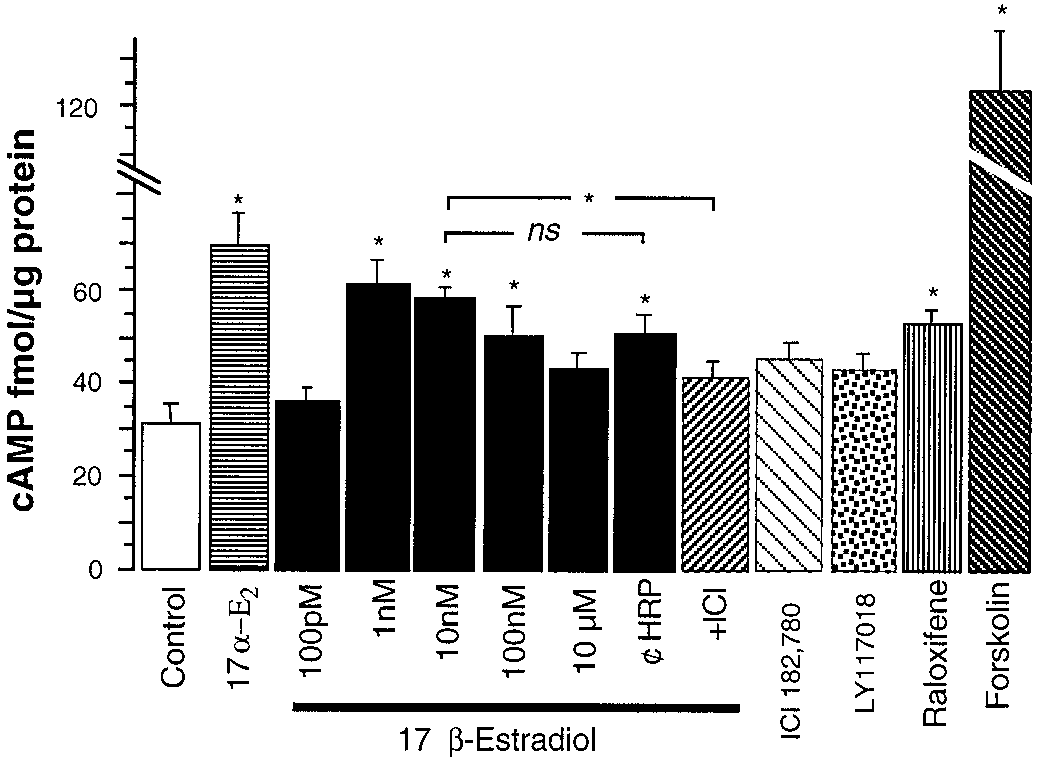
To explain the action mechanism of estrogens on exocytotickinetics, we analyzed the cell production of cAMP. Theseeffects are resumed in Fig. 7. The natural isomer 17-estra-diol increased cAMP only in the range of concentrations from1 to 100 nM. However, no significant change was observed at10 M. This effect of estrogen seems to be membrane-delim-ited as 17-estradiol HRP-conjugated also increased cAMP. No significant differences were observed between free andconjugated drug. The preincubation of cells with ICI 182,780
Fig. 6. The estrogen modulator LY117018 slowed down the time course of
abolished the effect of 17-estradiol on cAMP production.
exocytosis. Spikes were plotted incorporating the average values of the kinetic
Even when some estrogens produced changes in cAMP
parameters obtained from real data. Note the different ascending slope, the
production that were also observed in the time course of
and tP, as well as a decrease in the CA concentration
reaching the electrode. Net granule content of CA was, however, reduced.
secretory spikes, this was not a general rule. Hence, 17␣-estradiol, which was revealed to be inactive on secretion,
effects on “m ” did not exhibit a clear concentration depen-
increases the cAMP. Conversely, the LY117018 did not alter
dence, and full effects could be obtained with 10 nM (Table 2).
the cAMP levels but drastically affected exocytosis at the
Similar results were obtained with 17-estradiol and with
LY117018 (Table 2 and Fig. 5). Conversely, 17␣-estradiol did
Note, however, that the cAMP rise over the basal level was
not produce changes in the kinetics of exocytosis nor in gran-
modest when compared with 10 M forskolin. It was only
ule content. Surprisingly, the pharmacological profile of
evident after 15 min of drug incubation and required the
LY117018 was different than the other drugs tested, because
phosphodiesterase blockade with IBMX.
it caused a reduction to 79% in the apparent charge of gran-ules from 1.12 Ϯ 0.1 to 0.85 Ϯ 0.1 picocoulombs (Fig. 6),suggesting that this drug operates through an additional
Discussion
We are far from understanding which are the membrane
Estrogen Antagonist ICI 182,780 Also Exhibited Es-
targets for estrogens despite the wide number of nongenomic
trogen Activity. The compound ICI 182,780 has been pro-
effects of estrogens that have been described to date.
posed as a “pure estrogen antagonist” on the classical nuclear
In the present study, we combined secretory experiments
estrogen receptor. It means that its effects only will be evi-
on perfused adrenals with experiments of exocytosis on iso-
dent in the presence of an agonist. However, on exocytotic
lated cells to explore responses in these different prepara-
kinetics it seemed that ICI 182,780, at concentrations of 1 nM
tions as well as species differences. It was not only intended
or lower, behaved as an antagonist blocking estrogen action.
for two different species and preparations but also because
However, over this concentration it exhibited estrogenic ac-
culture maneuvers could cause alterations in the responses
tivity. Figure 5 shows the effects of ICI 182,780 on I
of a tissue like adrenal medullae presumably exposed to highlevels of steroids. However, the degree of inhibition observedin secretion was similar in both preparations, suggestingthat estrogenic transduction mechanisms were not highlyaffected by the isolation/culture processes.
Our results on perfused rat adrenal gland confirm and
extend previous results, which showed that acute applicationof estrogens caused the inhibition of secretory responses inchromaffin tissues when they were applied at micromolarconcentrations (Lo´pez et al., 1991; Park et al., 1996; Dar andZinder, 1997). Although the current study was not conductedto examine the target of estrogens to inhibit secretion, itseems that they affected the electrical properties of cell mem-brane. It could have occurred either by direct reduction ofCa2ϩ currents (Kim et al., 2000) or by increasing the Kϩconductivity (Minami et al., 1990; Valverde et al., 1999). Recently, Uki et al. (1999) described the inhibition of nico-tinic currents with high estrogen concentrations in rat cer-vical superior ganglia neurons. However, in chromaffin tis-
Fig. 7. Estrogens increased the intracellular cAMP levels. Cells were pretreated with 500
sue it is unlikely that estrogens act on neither the nicotinic
M IBMX for 15 min prior to another 15-min incu-
bation with Krebs’ solution (Control), 17␣-estradiol (17␣-E ), 17-estra-
receptor, because of the similar degree of inhibition observed
diol (at the indicated concentrations), 10 nM HRP-conjugated 17-estra-
between ACh and Kϩ, nor the secretory machinery, because
diol (¢-HRP), 10 nM 17-estradiol ϩ 1 nM ICI 182,780 (ϩICI), 1 nM ICI
LY117018 did not inhibit the Ca2ϩ-evoked secretion on
182,780, 10 nM raloxifene, 10 nM LY117018, or 1 M forskolin. Datawere the average of six different experiments. p Ͻ 0.05 (Tukey’s test).
A-23187-treated glands. The onset of the inhibition of CA
Machado et al.
secretion was rapid both in perfused glands and in isolated
isomer is inactive on classical receptors but inhibits secretion
bovine cells. The effect of 10 M estrogen was not affected by
in cat adrenals (Lo´pez et al., 1991).
a previous 1 M incubation, reinforcing the idea of the non-
One effect that was difficult to explain was the behavior of
“classical” antagonists on some nongenomic estrogen re-
The inhibition caused by estrogens on elicited [Ca2ϩ] was
sponses. In our hands, the ICI 182,780 antagonized the ef-
only observed at micromolar concentrations of estrogens as
fects of 17-estradiol on cAMP production (Fig. 7) and on the
occurred with the secretion of CA (Figs. 2 and 4).
kinetics of exocytosis (Fig. 5). However, it behaved as an
The most important observation of this study was, how-
agonist when applied alone (Table 2). This latter observation
ever, the effects of estrogens and related compounds at the
was in agreement with Ruehlmann et al. (1998) who also
level of single exocytotic events. This action of estrogens was
found that acutely administered ICI 182,780 mimicked the
fully observed at nanomolar concentrations and occurred
inhibitory effects of estrogens on Ca2ϩ currents of vascular
within seconds (Fig. 6 and Table 2). To our knowledge, this is
smooth muscle. The best explanation could be that this com-
the first description of the role of estrogens in the regulation
pound acts as a partial agonist with a higher activity but
lower intrinsic activity than estradiol.
It is not easy to address these effects of estrogens on
The pharmacological profile of estrogen modulators is still
exocytosis to an action mechanism. It can be discarded as a
far from being understood, and several differences have been
direct cytoplasmic effect of estrogens because the results with
found between tamoxifen and raloxifene. The compound
17-estradiol were reproduced using HRP-conjugated 17-
LY117018 also exhibited a different profile than raloxifene; it
estradiol. In addition, the use of nanomolar estrogen concen-
was more potent inhibiting secretion and slowing secretion,
trations makes improbable a direct effect on fluidity of lipid
but it did not increase cAMP levels. These results prevent us
membranes. Also, these effects occurred within seconds.
from attributing all of the membrane-mediated effects of
Therefore, estrogens should act on a membrane acceptor(s).
A few integral membrane proteins have been proposed as
reflects the concentration of CA reaching the
electrode. The distance between the surface of a carbon fiber
putative membrane receptors for estrogens, which include
electrode and the cell membrane is similar to the synaptic
the classical ␣-receptor expressed on the plasmalemma
cleft (Ϸ20 nM). It means that the kinetics of exocytosis could
(Razandi et al., 2000; Wyckoff et al., 2001). Another possible
control the concentration of neurotransmitter reaching the
target might be the -subunit of the maxi-Kϩ channels,
postsynaptic cell using the same vesicle content. It suggests
which directly produces hyperpolarization in the muscular
a new role for estrogens in the control of synaptic perfor-
cells of blood vessels (Valverde et al., 1999). However, we
mance. Chromaffin granules and dense cored vesicles, found
cannot find its consistent connection with the exocytosis.
in noradrenergic and other synapses, are similar organelles
Recently, Nadal et al. (2000) showed that estrogens could act
(Winkler and Fisher-Colbrie, 1998). Estrogen can be contin-
through the atypical ␥-adrenoreceptor as a “nonclassical ␣-
uously modulating the sympathetic nerve terminals like ar-
nor -estrogen receptor”; however, the presence of these re-
teriolar nerve-muscle synapses. We hypothesize that part of
ceptors has not been demonstrated so far in adrenomedullary
the protective actions of estrogens on vascular diseases in
premenopausal women could be mediated by this mecha-
We have recently found that second messengers like cGMP
(Machado et al., 2000) or cAMP (Machado et al., 2001) neg-atively modulated the kinetics of exocytosis. In the latter
Acknowledgments
article, we found that even a very modest rise of intracellularcAMP slowed down exocytosis whereas strong elevations,
We thank Dr. Rafael Alonso for the use of facilities to perform the
cAMP measurements and Antonio G. Garcı´a (Universidad Auto´noma
like forskolin treatment, also caused an increase in the net
de Madrid) for help with discussion of the manuscript. We are also
granule content of CA (Machado et al., 2001). In other words,
grateful to the personnel of the Matadero Insular de Tenerife for
both cGMP and low cAMP concentrations caused the decel-
their kind supply of cow adrenal glands. Carbon fiber to make elec-
eration of exocytosis. Both cyclic nucleotides are usually in-
trodes was the kind gift of Professor R. M. Wightman (University of
versely regulated (Soderling and Beavo, 2000). In addition to
the increase in cAMP production, acute treatment with es-trogens increased cGMP levels (and cGMP-dependent pro-
References
tein kinase activation) in pancreatic -cells (Ropero et al.,
Alvarez C, Lorenzo C, and Borges R (1997) Interaction between G protein-operated
receptors eliciting secretion in rat adrenals: a possible role of protein kinase C.
1999). This overlapping action of both second messengers
Biochem Pharmacol 53:317–325.
could explain the biphasic effect of 17-estradiol observed on
Anton AH and Sayre DF (1962) A study of the factors affecting aluminum oxide
trihydroxindole procedure for the analysis of catecholamines. J Pharmacol Exp
intracellular cAMP levels (Fig. 7), whereas the effects on
Ther 138:360 –375.
exocytotic kinetics remained almost constant in a wide range
Borges R (1993) Ionic mechanisms involved in the secretory effects of histamine in
the rat adrenal medulla. Eur J Pharmacol 241:189 –194.
of estrogen concentrations (Table 2). It is difficult to test the
Borges R, Travis ER, Hochstetler SE, and Wightman RM (1997) Effects of external
role of cGMP on type 2 phosphodiesterase in chromaffin cells
osmotic pressure on vesicular secretion from bovine medullary cells. J Biol Chem 272:8325– 8331.
because the low levels of both compounds obliged the use of
Chen ZJ, YU L, and Chang CH (1998) Stimulation of membrane-bound guanylate
IBMX, which strongly inhibits all phosphodiesterases.
cyclase activity by 17--estradiol. Biochem Biophys Res Commun 252:639 – 642.
Colliver TL, Hess EJ, Pothos EN, Sulzer D, and Ewing AG (2000) Quantitative and
Future investigations on nongenomic actions of estrogens
statistical analysis of the shape of amperometric spikes recorded from two popu-
should explain the different pharmacological profiles of
lations of cells. J Neurochem 74:1086 –1097.
Dar DE and Zinder O (1997) Short term effect of steroids on catecholamine secretion
LY117018 and raloxifene as well as why 17␣-estradiol be-
from bovine adrenal medulla chromaffin cells. Neuropharmacology 36:1783–1788.
comes inactive inhibiting rat adrenal secretion and the ki-
Falkenstein E, Norman AW, and Wehling M (2000a) Mannheim classification of
non-genomically initiated (rapid) steroid action(s). J Clin Endocrinol Metab 85:
netics of exocytosis but increases cAMP production. This
Estrogens and Exocytosis of Catecholamines
Falkenstein E, Tillmann HC, Christ M, Feuring M, and Wehling M (2000b) Multiple
genomic actions of 17-oestradiol in mouse pancreatic -cells are mediated by a
actions of steroid hormones: a focus on rapid, nongenomic effects. Pharmacol Rev
cGMP-dependent protein kinase. J Physiol (Lond) 521:397– 407. 52:513–555.
Ruehlmann DO, Steinert JR, Valverde MA, Jacob R, and Mann GE (1998) Environ-
Favit A, Fiore L, Nicoletti F, and Canonico PL (1991) Estrogen modulates stimula-
mental estrogenic pollutants induce acute vascular relaxation by inhibiting L-type
tion of inositol phospholipid hydrolysis by norepinephrine in rat brain slices. Brain
Ca2ϩ channels in smooth muscle cells. FASEB J 12:613– 619. Res 555:65– 69.
Segura F, Brioso MA, Go´mez JF, Machado JD, and Borges R (2000) Automatic
Gu Q and Moss RL (1996) 17-estradiol potentiates kainate-induced currents via
analysis for amperometrical recordings of exocytosis. J Neurosci Methods 103:151–
activation of the cAMP cascade. J Neurosci 16:3620 –3629.
Kawagoe KT, Zimmerman JB, and Wightman RM (1993) Principles of voltammetry
Shears SB (1991) Regulation of the metabolism of 1,2-diacylglycerols and inositol
and microelectrode surface states. J Neurosci Methods 48:225–240.
phosphates that respond to receptor activation. Pharmacol Ther 49:79 –104.
Kim YJ, Hur EM, Park TJ, and Kim KT (2000) Nongenomic inhibition of catechol-
Soderling SH and Beavo JA (2000) Regulation of cAMP and cGMP signaling: new
amine secretion by 17--estradiol in PC12 cells. J Neurochem 74:2490 –2496.
phosphodiesterases and new functions. Curr Opin Cell Biol 12:174 –179.
Tabares L, Ale´s E, Lindau M, and de Toledo GA (2001) Exocytosis of catecholamine
Lo´pez MG, Abad F, Sancho C, De Pascual R, Borges R, Maroto R, Dixon W, and
(ca)-containing and ca-free granules in chromaffin cells. J Biol Chem 276:39974 –
Garcı´a AG (1991) Membrane-mediated effects of the steroid 17-␣-estradiol on
adrenal catecholamine release. J Pharmacol Exp Ther 259:259 –285.
Uki M, Nabekura J, and Akaike N (1999) Suppression of the nicotinic acetylcholine
Machado JD, Morales MA, Go´mez JF, and Borges R (2001) cAMP modulates exocy-
response in rat superior cervical ganglionic neurons by steroids. J Neurochem
totic kinetics and increases quantal size in chromaffin cells. Mol Pharmacol72:808 – 814. 60:514 –520.
Valverde MA, Rojas P, Amigo J, Cosmelli D, Orio P, Bahamonde MI, Mann GE,
Machado JD, Segura F, Brioso MA, and Borges R (2000) Nitric oxide modulates a late
Vergara C, and Latorre R (1999) Acute activation of Maxi-K channels (hSlo) by
step of exocytosis. J Biol Chem 275:20274 –20279.
estradiol binding to the  subunit. Science (Wash DC) 285:1229 –1231.
Minami T, Oomura Y, Nabekura J, and Fukuda A (1990) 17 -estradiol depolariza-
Watson CS, Norfleet AM, Papas TC, and Gametchu B (1999) Rapid actions of
tion of hypothalamic neurons is mediated by cyclic AMP. Brain Res 519:301–307.
estrogens in GH3/B6 pituitary tumor cells via a plasma membrane version of
Moro MA, Lo´pez MG, Gandı´a L, Michelena P, and Garcı´a AG (1990) Separation and
estrogen receptor-alpha. Steroids 64:5–13.
culture of living adrenaline containing and noradrenaline-containing cells from
Wehling M (1997) Specific non-genomic actions of steroid hormones. Annu Rev
bovine adrenal medullae. Anal Biochem 185:185243–185248. Physiol 59:365–393.
Nadal A, Ropero AB, Laribi O, Maillet M, Fuentes E, and Soria B (2000) Nongenomic
Winkler H and Fisher-Colbrie R (1998) Regulation of biosynthesis of large dense-core
actions of estrogens and xenestrogens by binding at a plasma membrane receptor
vesicles in chromaffin cells and neurons. Cell Mol Neurobiol 18:193–209.
unrelated to estrogen receptor ␣ and estrogen receptor . Proc Natl Acad Sci USA
Wyckoff MH, Chambliss KL, Mineo C, Yuhanna IS, Mendelsohn ME, Mumby SM,
97:11603–11608.
and Shaul PW (2001) Plasma membrane estrogen receptors are coupled to endo-
Park YH, Cho GS, Cho ET, Park YK, Lee MJ, Chung JY, Hong SP, Lee JJ, Jang Y,
thelial nitric-oxide synthase through G␣i. J Biol Chem 276:27071–27076.
Yoo HJ, Choi CH, and Lim DY (1996) Influence of 17␣-estradiol on catecholamine secretion from the perfused rat adrenal gland. Korean J Intern Med 11:25–39. Address correspondence to: Dr. Ricardo Borges, Unidad de Farmacologı´a,
Razandi M, Pedram A, and Levin E (2000) Plasma membrane estrogen receptors
Facultad de Medicina, Universidad de La Laguna, E-38071 La Laguna, Tene-
signal to antiapoptosis in breast cancer. Mol Endocrinol 14:1434 –1447.
Ropero AB, Fuentes E, Rovira JM, Ripoll C, Soria B, and Nadal A (1999) Non-
Program Notes on Britten’s “Jubilate Deo” If you visit Baltimore, you should take time out from the crabcakes to visit some of the city’s religious sites, including the highest of all Anglo-Catholic parishes, Grace & St. Peter’s, the Roman Catholic Basilica, oldest in the United States, and the Museum of Visionary Art. Christopher Smart, author of Jubilate Agno, from which Ben
Blutfette – Zuckerkrankheit und Mitochondrien Seit Jahren waren bei Herrn M. hohe Cholesterin-Triglycerid- und Blutzuckerwerte bekannt. Fett- und cholesterinarme Diätregimes sowie hausärztliche Medikations-versuche verhinderten nicht, dass bis zum September 2005 die Cholesterinwerte auf fas 1000 mg/dl, die Triglyceride auf über 9000 mg/dl und der Blutzucker auf knapp 300 mg/dl anstiege
 Machado et al.
Machado et al.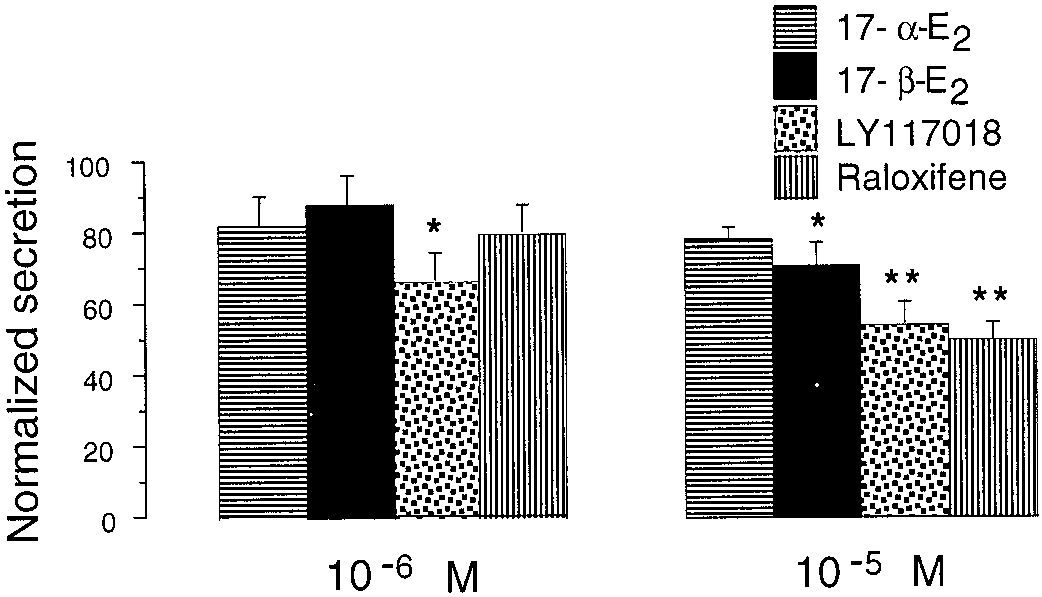
 Estrogens and Exocytosis of Catecholamines
Estrogens and Exocytosis of Catecholamines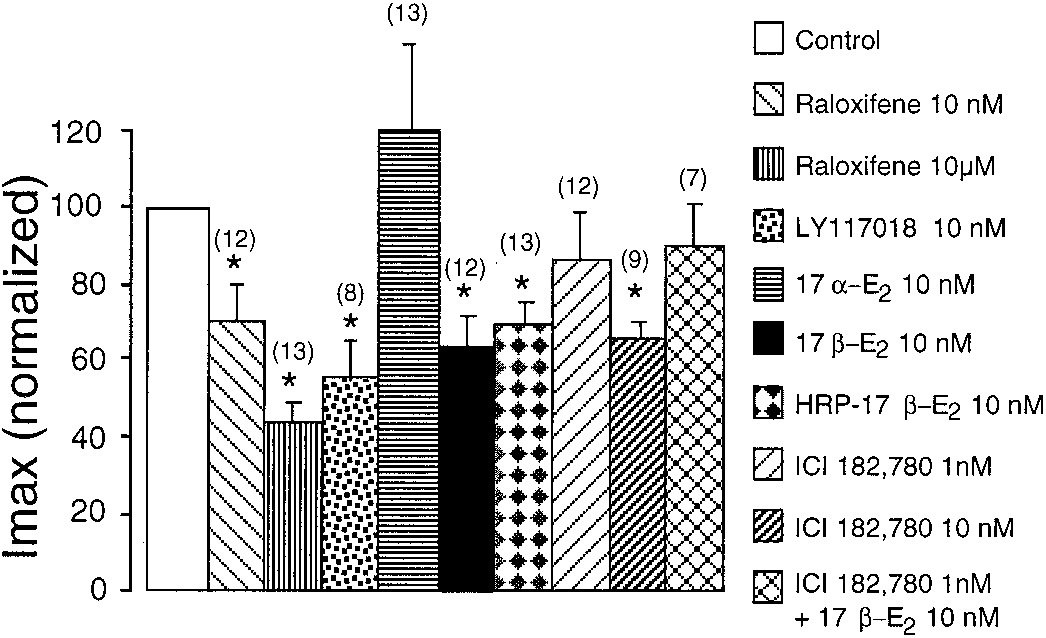

 Machado et al.
Machado et al.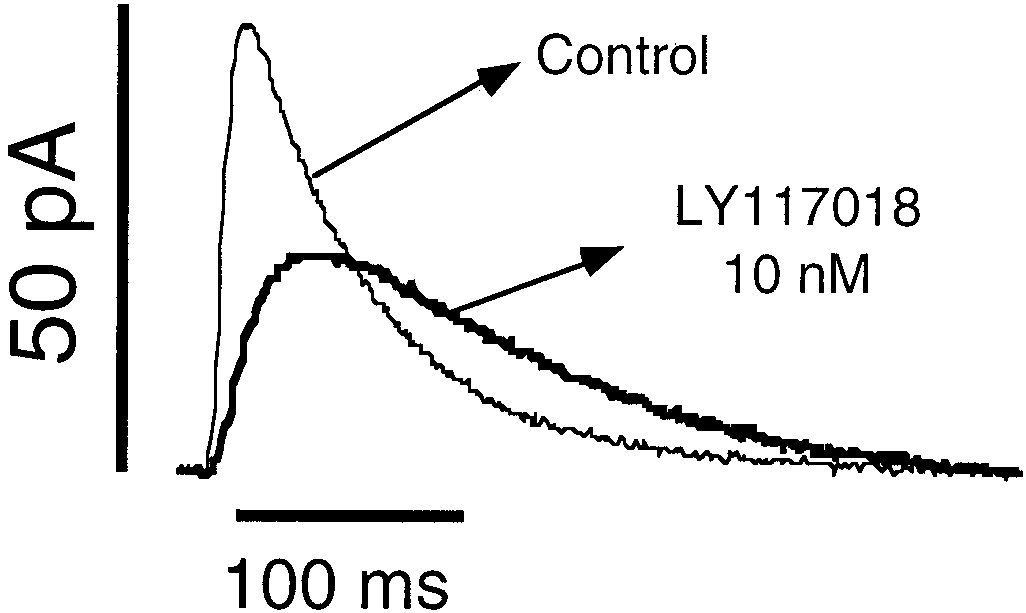
 Estrogens and Exocytosis of Catecholamines
Estrogens and Exocytosis of Catecholamines