Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
C2.013-22
Journal of Coastal Conservation 2: 13-22, 1996 EUCC; Opulus Press Uppsala. Printed in Sweden- Coastal sand dune vegetation: an extreme case of species invasion -Coastal sand dune vegetation: an extreme case of species invasion Castillo, S.A.1 & Moreno-Casasola, P.2*
1Laboratorio de Ecología, Facultad de Ciencias, UNAM, México 04510 D.F., México;
2Instituto de Ecología, A.C. Apdo. Postal 63, Xalapa 91000, Veracruz, México;*Fax + 52 28 186910 ext 1202; E-mail: patricmo@sun.ieco.conacyt.mxAbstract. The coastal sand-dune flora of the Gulf and Carib-
terized by a sandy, nutrient-poor substrate of siliceous
bean region of Mexico was analyzed to understand differ-
particles with variable amounts of calcareous sand. There
ences in floristic composition and richness found along the
are systems with narrow beaches and one or more paral-
coast. Each of the 655 species reported was classified accord-
lel dune ridges 3-8 m high. There are also topographically
ing to its ecology and distribution range by checking herbaria
complex systems with 25-m high dunes with different
specimens, literature and specialists. Three groups were
degrees of stabilization. Caribbean dunes in Mexico are
formed: (a) species with predominantly coastal distribution;(b) ruderal or secondary species frequently found inland,
formed by ridges of calcareous sand, derived from coral
common of disturbed areas such as roadsides, abandoned
reefs and shells as well as from the limestone Peninsula
fields or forming part of secondary growths; (c) inland spe-
of Yucatan. Both regions have many floristic and
cies frequently found in other vegetation types such as tropi-
physiognomic differences (Sauer 1967; García 1987;
cal dry or seasonal forest and grassland. A total of 71 coastal
species, 237 ruderal/secondary and 336 species from other
In patch dynamics, natural systems are seen as mo-
community types were found. The distribution of these groups
saics of local, suitable patches colonized by various
was analyzed along 44 sites of the Gulf and Caribbean, in the
species at different times. Species ranges show that a
different dune habitats and for the dominant growth forms.
species’ internal geographical abundance is usually dy-
Coastal species are more widely distributed; they predomi-nate in habitats with sand movement and the herbaceous
namic, even when the range limits are stable. Thus,
component prevails. Ruderal/secondary species and especially
invasions can be conceived as temporary disturbances
those belonging to other vegetation types frequently appear in
leading to spatial dynamism and viewed more broadly,
only one or two sites occupying more protected or stabilized
species ranges can be regarded as being in a constant
habitats. The two latter groups considerably increase species
state of flux, both internally and externally (Hengeveld
richness of sand dune flora, but also pose interesting prob-
1989). Plant community boundaries are not static;
ecotones show a constant exchange of species. Seeddispersion can take place between communities that are
Keywords: Coastal dune; Species distribution; Richness.
Invading species have been identified as those being
indigenous to some region other than the area being
Introduction
invaded. In many cases, this is not always clear-cut: it isdifficult to define community boundaries, modification
The floristic composition of a plant community is
of scale in the analysis makes results differ, and popula-
the result of its disturbance history, its present dynam-
tion density as well as species presence should be taken
ics, the life history (attributes) of the species colonizing
into account to define the native range of a species. For
and reproducing successfully in that particular environ-
the purpose of this paper a difference is made between
ment and species availability (Guevara 1982). Sand
exotics and invaders. Exotics can be defined as alien
dune vegetation is a clear example of this and we believe
species introduced intentionally or non intentionally to
that invasion of species with different traits are impor-
the region. Often, ecosystem invasion by exotic plants
tant contributors to floristic richness and biodiversity.
can have detrimental effects. Exotics displace native
Once their propagules have dispersed into the dune
species by competing for resources, interfere with suc-
systems, species attributes (such as germination, estab-
cessional processes, alter disturbance regimes and dis-
lishment and growth requirements) will be important in
rupt food chains (Van Wilgen & Richardson 1985;
defining in which part of the dune mosaic invading
MacDonald & Frame 1988; Elliott & White 1989). We
define invaders as species with an occasional presence
Coastal sand dunes in the Gulf of Mexico are charac-
for that type of community (arbitrary frequency values
Castillo, S.A. & Moreno-Casasola, P.
of 10-20% for the sites) which can have low or high
nity invasibility varies according to the characteristics
population numbers on a site and are frequently charac-
of the community and the life history traits of the po-
teristic of other community types. Huston (1994) con-
tential invaders (Ewel 1986). Disturbed communities
siders that invasions potentially lead to an increase in
are more often subject to invasions than intact commu-
species richness, as the invading species are added to the
nities and disturbance regime (type, frequency, dura-
existing species pool, although numerous examples dem-
tion and magnitude of disturbance) is recognized as the
onstrate that they can also lead to extinctions.
most important overall factor allowing invasion of ex-
In biological invasions there are two components,
otic species (Elton 1958; Myers 1984; Ewel 1986; Fox
modulated by timing and chance: the invasive species
& Fox 1986; Crawley 1987; Rejmánek 1989; DeFerrari
and the invaded community. Consequently, commu-
& Naiman 1994; Huston 1994). However, there are
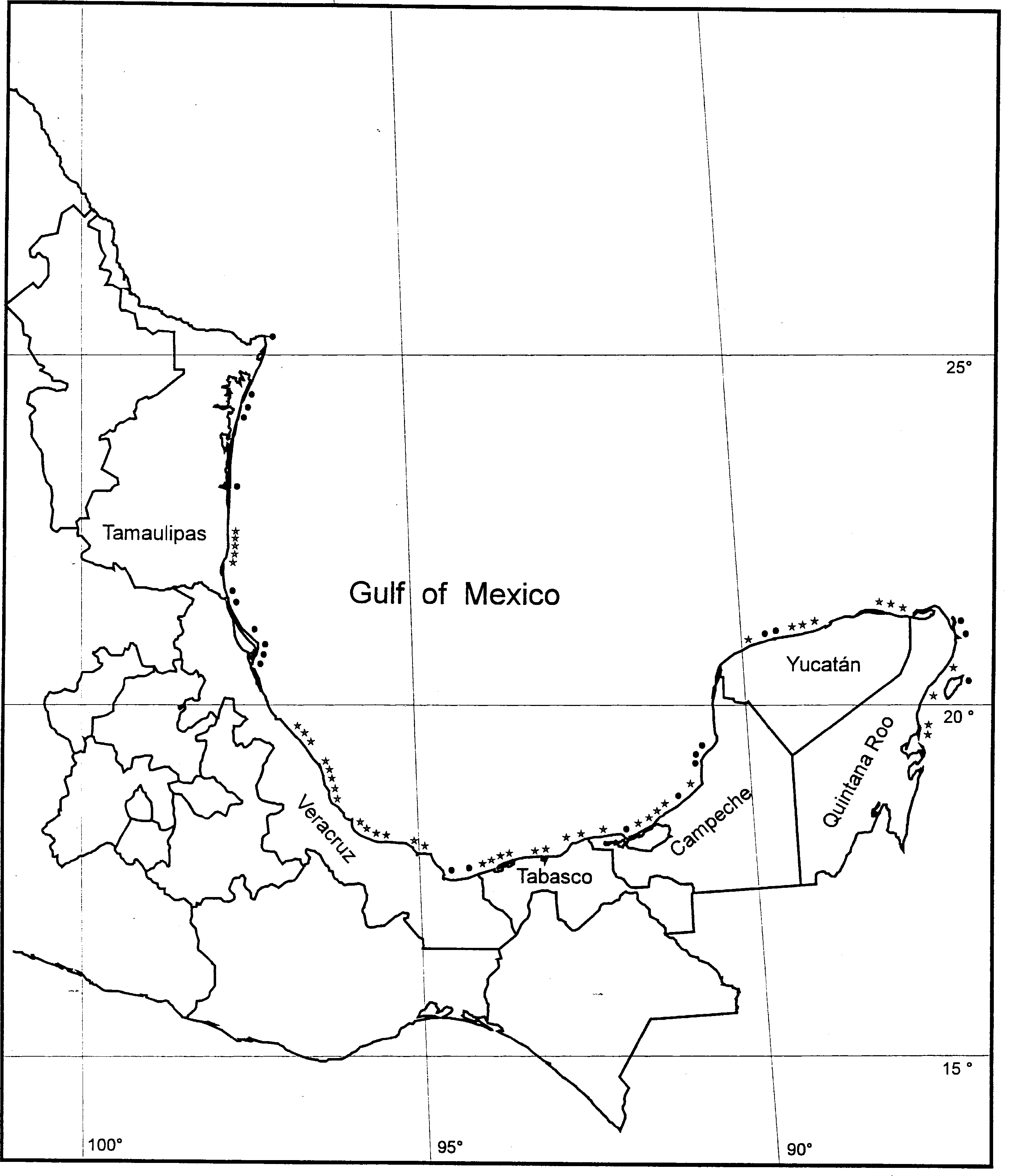
Fig. 1. Location of the 44 beach and dune study sites sampled (*). Sites include the Gulf (Northern Gulf- sites 1 to 6 and Central Gulf- sites 7 to 29) and the Caribbean (sites 30 to 44). Other beaches and dunes studies (reported in the literature) are indicated with a dot. See text for references. - Coastal sand dune vegetation: an extreme case of species invasion -
several examples in which invasion occurs in the ab-
several species mainly because of absence of herbaria
sence of disturbance (Mack 1985; Burdon & Chilvers
collections and these were left out of the calculations
1977; Kruger 1977). Huston (1994) considers that there
(Amaranthus arenicola, Eragrostis excelsa, E. yucatana,
has been little progress in generalizations about which
Eriochloa boxiana, Mandevilla subsagittata, Matelea
ecosystems are most likely to be invaded. yucatanensis, Panicum rigidulum, Paspalum clavu-
Coastal dunes are very dynamic systems comprising
liferum, Pithecellobium grisebachi, Solanum yucata-
a wide variety of habitats with different physical and
biotic conditions, allowing the existence of species with
Species distribution patterns were divided into three
very diverse life history traits. They can be visualized as
a permanently changing environment with distinct de-
(1) species with a predominantly coastal distribution
grees of stabilization closely correlated with the topog-
(sand dunes, coastal marshes or mangroves) henceforth
raphy and the disturbance produced by sand movement
(Barbour et al. 1985; Moreno-Casasola 1986; Maun &
(2) inland ruderal or secondary species frequently found
Lapierre 1984) and slack inundation (van der Laan
inland and common inhabitants of disturbed areas such
1979; Sykes & Wilson 1987; Grootjans et al. 1991). It
as roadsides, abandoned fields or secondary growths;
results in a vegetation mosaic which has been described
(3) inland species, frequently found in other vegetation
The aim of this work is to understand the floristic
types (OT) which can be considered mature, such as
composition, richness and variability of the regional
tropical seasonal forest, oak woodland or grassland.
sand-dune systems along the Gulf and Caribbean through
For most of the analyses the complete floristic list
a detailed analysis of species composition, trends in
was used (655 species). Where geographical or local
species distribution patterns, species richness under dif-
habitat distributions were analyzed only data for the 44
ferent habitats and disturbance regimes and growth forms
sites sampled with the same methodology and intensity
that compose the present dune flora. Strategies for con-
were utilized (492 species), so as to avoid errors caused
servation and management of the dune flora will be
by intensity of sampling during floristic collections.
influenced by the resulting species analysis.
Species numbers varied considerably among sites
We accumulated a list of 655 species for the coastal
and distribution patterns. A total of 71 (10.83%) coastal
beach and dune vegetation along the Gulf and Carib-
species (C) were found. The sand dune flora also com-
bean Sea of Mexico. A data base was elaborated to
prised 237 ruderal/ secondary species (R/S- 36.2 %), as
allow for consultations. Data from the vegetation analy-
well as 336 species characteristic of other communities
sis of 44 beach and dune sites (Fig. 1) totalling 492
(OT- 51.3 %). Species richness per site (Fig. 2) is very
species was used (Moreno-Casasola et al. 1982; Espejel
variable (18 - 157 species). The number of species in the
1984; Moreno-Casasola & Espejel 1986; García 1987;
three categories (out of a total of 492) in each of the 44
Castillo et al. 1991). This list was augmented with
sites also varies considerably. The mean number of C-
information from publications (Flores 1984; Gonzalez-
species found was 15.5 (± 4.6), of R/S-species was 18.5
Medrano 1972; Poggie 1962; Puig 1976; Sauer 1967)
(± 9.7) and OT-species was 21.4 (± 14.7).
and herbaria specimens in the National Herbarium at the
Although the coastal flora between the Gulf and the
National Autonomous University of Mexico (MEXU)
Caribbean is quite distinct in species numbers and com-
and in the Herbarium of the Institute of Ecology in
position (Sauer 1967; Moreno-Casasola & Espejel 1986;
Xalapa (XAL). Finally, a partial list of some of the
Espejel 1984; Moreno-Casasola 1988) the same trends
species belonging to each species type is given in Table
in species number per pattern of distribution is main-
4. The complete list is available from the authors.
tained (Table 1). To further understand this distribution,
Each of the 655 species was assigned to one of
a more detailed analysis of the flora of the Gulf coast
three categories of distribution patterns. This was done
was made by subdividing this region’s flora into two
by examining all specimens for each of the listed spe-
distinct floristic groups based on distribution of pre-
cies in the collections of both herbaria. A category was
dominant growth forms, presence of endemics, biogeo-
assigned when 60% or more of the specimens indi-
graphical relations and general floristic composition
cated a certain type of distribution. This was further
(Moreno-Casasola 1988, 1991). The Northern Gulf group
checked against the literature and by specialists (see
(State of Tamaulipas and northern part of Veracruz –
Acknowledgements). We were not able to classify
sites 1 to 6 in Fig. 2 – has many similarities with the
Castillo, S.A. & Moreno-Casasola, P.Fig. 2. Species richness along the 44 Gulf and Caribbean sites. Black low parts of columns represent coastal species (C), open middle parts species from other vegetation types (OT), and shaded parts ruderal/ secondary species (R/S).
Texas coastal flora; the Central Gulf group (States of
higher frequency values. This means that more species
Veracruz and Tabasco – sites 7-29) has many different
are found along a high number of beach and dune sites.
elements. Comparisons between both Gulf groups show
The species most frequently occurring on the beach and
that the Central group has the lowest proportion of C-
embryo dunes along the Gulf coast are: Ipomoea
species, sharing 32 species with the Northern group. stolonifera, Oenothera drummondii, Chamaecrista
Species numbers vary between floristic groups but pro-
chamaecristoides, Schizachyrium scoparium var.
portions of the three categories are roughly maintained,
littoralis, Palafoxia lindenii, Amaranthus greggii and
especially when comparing both Gulf groups with the
Fimbristylis spadicea. They are present in more than
half of the sampled sites. Several of them are important
Species presence along sites varied greatly. A high
sand stabilizers in active dunes, which are found along
number of species were very infrequent, appearing only
this coast. Hibiscus pernambucensis is less frequent but
in one or two sites. Species frequency distribution was
also a characteristic tree species along these coasts.
analyzed for the three categories. Fig. 3a, b shows that
Another group of beach species is mainly found along
C-species, both along the Gulf and the Caribbean, have
the Caribbean (Scaevola plumierii, Coccoloba uvifera,Table 1. Number of species (out of a total of 655) and percentage of coastal (C), ruderal/secondary (R/S) and species from other vegetation types (OT) found in the Gulf and Caribbean regions. The Northern Gulf floristic group is found in Tamaulipas and North Veracruz, the Central Gulf group inhabits central and southern Veracruz and Tabasco and the Caribbean group includes Campeche and the Yucatan Peninsula (Moreno-Casasola 1991). - Coastal sand dune vegetation: an extreme case of species invasion -Fig. 3. Frequency distribution of coastal species (C) along 44 sites on the Gulf (a) and Caribbean (b). Frequency distribution of ruderal/secondary species (R/S) -shaded bars- and for species characteristic of other vegetation types (OT) -darkened columns- along the Gulf (c) and Caribbean (d). See text for explanation. Ambrosia hispida, Tournefortia gnaphalodes, Ernodea
of R/S and 62 % of OT-species were recorded in one or
littoralis, Suriana maritima, Tribulus cistoides) and still
two sites (in contrast with 52 % of C-species in seven
other species (Ipomoea pes-caprae, Sesuvium portula-
sites). Values on the Y-axis are much higher in Fig. 3b
castrum, Sporobolus virginicus, Canavalia rosea,
than in Fig. 3a. Along the Gulf, 43.2 % of R/S-species
Okenia hypogea, Croton punctatus) are widely distrib-
appeared in only 1-2 sites and only seven species (Bidens
uted in both areas. Coastal species in stabilized dunes
pilosa, Panicum maximum, Commelina erecta, Iresine
also vary between the Gulf and Caribbean. Schiza-diffusa, Lippia nodiflora, Porophyllum punctatum and
chyrium scoparium var. littoralis, Schrankia quadri-Crotalaria incana) occur in more than a third of the
valvis and Cenchrus tribuloides are frequent along the
sampled sites (which is equivalent to ten sites). For the
Gulf. In the Caribbean the following species are com-
Caribbean 68.5 % of the species appeared in 1-2 sites
mon: Tribulus cistoides, Ernodea littoralis, Scaevola
and seven species (Lantana involucrata, Jacquiniaplumierii, Coccoloba uvifera and Pithecellobiumaurantiaca, Gossypium hirsutum, Gymnopodium flori-keyense. The latter form dense thickets. bundum, Cenchrus echinatus, C. incertus and Por-
The frequency distribution of R/S and OT-species
tulaca pilosa) are also found in more than one third
follows a much more pronounced J-curve (Fig. 3c, d).
of the sites (seven sites). Passiflora foetida, Por-
Many of them occur in only one or two sites, and very
tulaca oleracea, Commelina erecta, Macroptilium
few appear in more than one third of the sites. Data for
atropurpureum, Malvaviscus arboreus and Rivina laevis
Gulf and Caribbean species together, showed that 55 %
are widely distributed in the Gulf and Caribbean. Castillo, S.A. & Moreno-Casasola, P.Table 2. Number and percentage of coastal species (C), ruderal/secondary (R/S) and species from other vegetation types (OT) in habitats of the dune system. Percentages in bold letters indicate values along horizontal lines. Three main habitats based on the subdivisions elaborated by Doing (1981) and Moreno-Casasola & Espejel (1986) are used. Dune habitats are characterized mainly by sand movement (and low salinity). Water-dependent habitats are the slacks where the elevation of the water table produces inundation during the rainy season and a higher concentration of nutrients. Stabilized areas — such as grasslands, thickets or tropical forest — have a dense plant cover, no sand movement, roots are out of reach of the phreatic table and physical factors lose importance as agents of disturbance.
OT-species show the same trend as R/S-species (Fig.
rises. OT-species also represent more than half of the
3c, d). Many of the Gulf species (60.9%) appear exclu-
species present (53%), R/S represent more than a third
sively in one or two sites and only a few appear in more
(34.5%) and C-species 12.3%. In stabilized habitats
than a third of them (Chamaesyce dioica, Cyperus articu-
where there is no sand movement and conditions are less
latus, Hydrocotyle bonariensis, Erigeron myrionactis,
stressful such as grassland, thickets and tropical forest
Chamaesyce ammanioides, Citharexyllum ellipticum,
— including the protected area behind the foredune —,
Cissus sicyoides and Psychotria erythrocarpa). Shared
OT-species represent more than half (54%), R/S-spe-
by the Gulf and Caribbean beaches and dunes are
cies 38.5% and C-species only 7.4%. If we analyse
Chiococca alba, Metastelma pringlei, Randia laete-
distribution of C-species among habitats (Table 2, per-
virens, Waltheria indica and Bursera simaruba. In the
centages in bold letters), we find that 41% of them occur
Caribbean 64.2% species occur in 1-2 sites and 11spe-
in dune habitats, 37.8% in stabilized communities and
cies appear in more than one third (Agave angustifolia,
21% in slacks or water-dependent habitats. R/S and OT-
Coccothrinax readii, Cordia sebestena, Capparis
species have low values in dune habitats (11 and 7.2%
flexuosa, C. incana, Commicarpus scandens, Metopium
respectively), intermediate values in slacks (20.5 and
brownei, Thrinax readii, Bumelia retusa, Caesalpinia
22.9% respectively) and high percentages in stabilized
vesicaria and Hymenocallis littoralis).
Coastal dune vegetation is characterized by a mosaic
Growth forms also vary among the species groups
of habitats in which environmental factors change. C, R/
(Table 3). Herbaceous species can be divided into (1)
S and OT-species differ in their presence among habi-
grasses and sedges — in which OT-species account for
tats (Table 2). Dune habitats are formed by the sandy
48.3% and R/S-species for 33.3% — and (2) forbs
beach, embryo dunes and foredunes in which sand move-
(where R/S-species account for 45.1%). OT-species are
ment is the dominating environmental factor. C-species
the dominant group among shrubs (51.2%), and espe-
predominate in the dune habitat (40.6%), R/S and OT-
cially trees, where they represent 77.6%. In both shrubs
species represent 31.21% and 28.1% respectively. Hu-
and trees coastal species have few representatives. If we
mid and wet slacks are those habitats which become
analyse the growth forms of C-species (Table 3, per-
inundated during the rainy season when the water table
centages in bold letters), we find that 71% of them are
Table 3. Number and percentage of coastal (C), ruderal/secondary (R/S) and species from other vegetation types (OT) for the various growth forms. Other growth forms include cacti, agaves and epiphytes. Percentages in bold letters indicate values along horizontal lines. - Coastal sand dune vegetation: an extreme case of species invasion -Table 4. Partial list of coastal (C), ruderal/secondary (R/S) and species from other vegetation types (OT). Ruderal / secondary Other vegetation types
herbaceous (grasses and sedges: Schizachyrium scopa-cia farnesiana, Celtis pallida, Caesalpinia cacalaco,rium, Panicum amarum, Sporobolus virginicus, Trachy-Guazuma ulmifolia, Senna occidentalis, Thevetiapogon gouini, Uniola paniculata, Fimbristylis spadicea,gaumeri, Verbesina persicifolia). OT-species show a
and forbs: Ipomoea pes-caprae, Canavalia rosea. See
different trend. Herbaceous elements represent 43.9%
Table 4). 27.5% of the C-species are woody (shrubs or
(Eleocharis spp., Fimbristylis castanea, Nymphaea
trees: Chrysobalanus icaco, Ernodea littoralis, Cocco-ampla, Typha domingensis, Rhynchosia americana)
loba uvifera, Chamaecrista chamaecristoides, Hibiscus
and woody species account for more than half (51.2%
pernambucensis, Suriana maritima) and 0.7% have par-
— Acacia cornigera, Bursera simaruba, Bumelia spp.,
ticular growth forms such as cacti (Opuntia stricta). Cedrela odorata, Enterolobium cyclocarpum). Palms
R/S-species are also mainly herbaceous (66.8% -
were considered under shrubs and trees, because of
Asclepias curassavica, Cenchrus echinatus, Commelina
their physiognomy (Acrocomia mexicana, Thrinaxerecta, Desmodium spp., Macroptilium atropurpureum,radiata, Coccothrinax readii, Scheelea liebmanii). No
Rhynchelitrum repens, Sida rhombifolia, Solanum
other data are available on various ecological or physi-
nigrum) and the woody species represent 33.1% (Aca-Castillo, S.A. & Moreno-Casasola, P.Discussion
Mexico. 46% of the taxa were perennial, non-succulentforbs, 29% were perennial grasses and allies and 26 %
Frequently, exotics are considered synonymous to
woody perennials, mostly sclerophyllous. Percentages
invaders. There are few data on dune colonization by
of herbaceous and woody C-species in dune habitats are
exotics from abroad (Rhynchelytrum repens, Cocos
very similar to those found by Barbour et al. (1987). nucifera, Casuarina equisetifolia). Exotics are known
Most R/S and OT-species were found in only one or
to compete with native species, even to the point of
two sites. They invade the dunes and some propagules
replacing them. It is difficult to imagine the implications
are able to germinate and establish. Stabilized habitats
that such invasions have had on dune systems, as studies
and slacks have adequate conditions for their successful
on dune communities are recent, and human activities
establishment and reproduction. Their presence prob-
both on the coastal plain and on dunes are quite old.
ably depends on nearby sources, time, chance, species
Johnson & Barbour (1990) mentioned three exotics that
attributes and adequate microhabitats in the dune sys-
tend to take over habitats they invade in the coasts of
tem. These are found further away from the beach:
Florida: Agave sisalana, Schinus terebinthifolius and
humid slacks and stabilized areas. OT-species are able
Casuarina equisetifolia. In this study invaders (OT and
to invade as well as S/R-species but with lower num-
R/S) are those species which have not evolved in coastal
bers. Slacks are also subjected to disturbances produced
environments, but their life history traits allow them to
by inundation. Wetland flora is adapted to these condi-
colonize and reproduce successfully. Their presence is
tions and easily invades new sites in which humid
highly variable and they are not so frequent as to be
conditions predominate. This would explain the high
considered characteristic elements of the communities.
number of OT-species found in water dependent habi-
59% of invaders are found in just one or two sites. The
tats. R/S-species are able to colonize newly opened
analysis performed on the 655 species that constitute the
spaces in which vegetation has died after a prolonged
coastal dune flora of the Gulf and Caribbean of Mexico
helps to understand why dune communities behind the
Species adapted to stable mature communities (OT)
foredune are so different both floristically and structur-
are constantly invading and colonizing. Succession tends
ally from one place to another. The dune system seems
towards grasslands and tropical forests — dry and
to tolerate and enrich its flora with invaders.
semideciduous forests — (Novelo 1978; Moreno-
The results of this study show that species richness
Casasola & Espejel 1986) where OT-species will domi-
for any one site depends on the presence of several
nate. They show the highest percentage of invaders
groups of species, each of which predominates in par-
(Table 2), having more OT-species than S/R species.
ticular habitats within the dune system and in different
The standard deviation for OT-species number along
periods of the stabilization process. There is essentially
the 44 sites is the highest (21 ±14.7), the minimum being
no biological difference between the process of invasion
6 and the maximum 184. In these habitats there is
and the process of colonization or recolonization by
usually an almost closed vegetation cover and probably
native plants. Colonization after a disturbance is a fun-
more competition among plants both for nutrients and
damental process of plant succession (Huston 1994). C-
water. Disturbances are more sporadic and less intense.
species, which include those solely adapted to sand
Thickets and tropical forests on coastal dunes are mainly
dune environments, mangroves and coastal marshes,
formed by invaders (mostly OT-species) and with time
have a more uniform distribution throughout the littoral
will resemble these types of communities established
areas (Fig. 2 and Table 1) although, numerically, they
inland, although always with some coastal elements (C-
often constitute a minor component on each site (Fig. 2).
Populations may disappear from any one area, but
The results obtained for the three habitat types do
they will soon recolonize and reestablish. They occupy
not show any direct relationship between disturbance
environments with stressful physical conditions (sand
and invasibility. R/S-species account for approximately
movement, airborne salinity, sea-water inundation) in
one third of the richness in the three of them, regardless
which other species are not successful such as beaches,
of their disturbance regime. OT-species account for
embryo dunes and active dunes. In these habitats distur-
54% of the flora of stabilized sites, where sand move-
bance by sand movement is determinant and is con-
ment and inundation are not important.
stantly opening new spaces for colonization. C-species
During the last decades, many tropical sand dune
are the most abundant here and they have probably
areas have increasingly been bordered by agricultural
evolved under these drastic conditions (Table 2). This
land. The coastal plain has rich soils with crops and
explains their higher frequency distribution along the
managed grasslands for cattle. They are connected by
sites (Fig. 2). Barbour et al. (1987) analyzed growth
dirt roads and other types of corridors that favour the
forms of beach vegetation along the northern Gulf of
spread and persistence of weeds, thus facilitating
- Coastal sand dune vegetation: an extreme case of species invasion -
movement of plants through the landscape (DeFerrari
and are associated with the first stages of dune
& Naiman 1994; Romano 1990). Human activities fa-
stabilization. Dune conservation should reproduce spa-
vour the presence of weeds and ruderal species, many
tially the temporal sequence of dune stabilization. By
of which produce small, easily dispersed seeds. It is
favouring this mosaic of habitats, species whose exist-
probable that the disappearance of forests and the in-
ence depend on the physical factors determining each
crease in croplands, pastures and secondary vegetation
habitat are guaranteed a place where they can reproduce
will increase the R/S component and modify the com-
successfully. Later stages favour the presence of invad-
position of the vegetation covering stabilized dunes.
ers (OT and R/S) which drastically increase local
This will increase the presence of herbaceous growth
biodiversity. Their presence probably depends on nearby
Many other physiological parameters will also be
The data presented in this paper seem to imply a
modified. As Barbour et al. (1985) have shown, beach
contradiction between favouring habitats for C-species
species show trends in life forms, leaf traits and physi-
and facilitating stabilized conditions more adequate for
ological behaviour (for example germination, growth
OT-species (and quite a number of R/S-species) and
and photosynthesis have broad temperature optima).
thus richer communities. We believe dune conservation
We do not have information on these differences for C,
strategies should take into account the simultaneous
maintenance of several degrees of stabilization in nearby
Currently, the result is a highly variable flora be-
systems as well as preserved patches of communities
tween different dune systems in a geographic region, the
inland (forests, thickets, grasslands) which can act as
coastal element (C) being the common link because of
seed sources. Efforts should be encouraged to preserve
its higher frequency distribution. At the regional level
the dynamics and ecosystem functioning of dunes, which
other vegetation types, i.e. mediterranean-type shrub-
include both the habitats in which coastal species sur-
lands, also show a high level of floristic variation al-
vive successfully and those in which thickets and trees
though structure remains fairly uniform (Griffin et al.
Biodiversity and its conservation is now becoming
one of the main concerns of humankind. Coastal dune
Acknowledgements. We are grateful to Jerzy Rzedowski,
systems occupy a very low percentage of the land
Gonzalo Castillo, Patricia Dávila, Fernando Chiang and Javier
environments. In subtropical and tropical regions it is
Valdés for checking the assigned species categories, and to
rapidly diminishing with the impressive growth of
Sergio Guevara, Carlos Vazquez-Yanes and Guadalupe
touristic developments. Species richness in dune sys-
Williams-Linera for their helpful comments on an earlier
tems cannot be approached in a simple way. It varies
geographically (Gulf and Caribbean) as well as locally(Figs. 1 and 2), depending on the degree of stabilization,mosaic of environmental conditions, floristic compo-
References
sition (percentage of C, OT and R/S-species) and po-tential sources of propagules of invading species. The
Barbour, M., DeJong, T.M. & Pavlik, B.M. 1985. Marine
beach and dune plant communities. In: Chabot, B.F. &
dune flora must be considered as a floristically vari-
Mooney, H.A. (eds.) Physiological ecology of North Ameri-
able and rich entity formed by species with very differ-
can plant communities, pp. 296-322. Chapman and Hall,
ent evolutionary histories. The sand dune patchwork of
environmental conditions and communities changes in
Barbour, M., Rejmánek, M., Johnson, A.F. & Pavlik, B.M.
space and also in time. It is a highly dynamic system in
1987. Beach vegetation and plant distribution patterns
which processes of succession and disturbance con-
along the northern Gulf of Mexico. Phytocoenologia 15:
stantly produce heterogeneity and allow the presence
Burdon, J.J. & Chilvers, G.A. 1977. Preliminary studies on a
Dunes are complex mosaics of habitats, each with
native Australian eucalypt forest invaded by exotic pines.
dynamics that are closely linked with particular distur-
Oecologia (Berl.) 31: 1-12.
Castillo, S., Popma, J. & Moreno-Casasola, P. 1991. Coastal
bance regimes (mainly sand movement and inundation).
sand dune vegetation of Tabasco and Campeche, Mexico.
Conservation and management of dune communities
should take into account the preservation of this hetero-
Crawley, M.J. 1987. What makes a community invasible? In:
geneity, as each element-type of this patchwork re-
Gray, A.J., Crawley, M.J. & Edwards, P.J. (eds.) Coloni-
sponds differently to human alterations. Of primary
zation, succession and stability, pp. 429-453. Blackwell
concern are habitats needed by C-species. These habi-
tats are characterized by the intensity of disturbances
DeFerrari, C.M. & Naiman, R.J. 1994. A multi-scale assess-
Castillo, S.A. & Moreno-Casasola, P.
ment of the occurrence of exotic plants on the Olympic
Moreno-Casasola, P. 1986. Sand movement as a factor in the
Peninsula, Washington. J. Veg. Sci. 5: 247-258.
distribution of plant communities in a coastal dune sys-
Doing, H. 1981. A comparative scheme of dry coastal dune
habitats, with examples from the eastern United States and
Moreno-Casasola, P. 1988. Patterns of plant species distribu-
some other temperate regions. Veröff. Geobot. Inst. Eidg.
tion on coastal dunes along the Gulf of Mexico. J. Biogr.Tech. Hochsch. Stift. RübelZür. 7: 41-72
Elliott, K.J. & White, A.S. 1989. Competitive effects of vari-
Moreno-Casasola, P. 1991. Sand dune studies on the eastern
ous grasses and forbs on ponderosa pine seedlings. For.
coast of Mexico. In: Davidson-Arnott, J. (ed.) Proc. Cana-dian Symposium on Coastal Sand Dunes. pp: 125-230,
Elton, C.S. 1958. The ecology of invasions by animals and
September 1990, Guelph, Ontario. National Research
Espejel, I. 1984. La vegetación de las dunas costeras de la
Moreno-Casasola, P. & Espejel, I. 1986. Classification and
Península de Yucatán. I. Análisis florístico del estado de
ordination of coastal sand dune vegetation along the Gulf
Yucatán. Biotica 9: 183-210.
and Caribbean Sea of Mexico. Vegetatio 66: 147-182.
Ewel, J. 1986. Invasibility: lessons from South Florida. In:
Moreno-Casasola, P., van der Maarel, E., Castillo, S., Huesca,
Mooney, H.A. & Drake, J.A. (eds.) Ecology of biological
M.L. & Pisanty, I. 1982. Ecología de la vegetación de
invasions of North America and Hawaii, pp. 214-230.
dunas costeras: estructura y composición en el Morro de la
Mancha I. Biotica 7: 491-526.
Flores, S. 1984. Vegetación insular de la Península de Yucatán.
Myers, R.L. 1984. Ecological compression of TaxodiumBol. Soc. Bot. Méx. 45: 23-37. distichum var. nutrans by Melaleuca quinquenervia in
Fox, M.D. & Fox, B.J. 1986. The susceptibility of natural
southern Florida. In: Ewel, K.C. & Odum, H.T. (eds.)
communities to invasions. In: Groves, R.H. & Burdon, J.J. Cypress Swamps. Univ. of Florida Press, Gainesville, FL.
(eds.) Ecology of biological invasions, pp. 57-66. Cam-
Novelo, R.A. 1978. La vegetación de la Estación Biológica El
Morro de La Mancha, Veracruz. Biotica 3: 9-23.
García, M.T. 1987. Descripción de la vegetación de dunas
Poggie, J.J. 1962. Coastal pioneer plants and habitat in the
costeras del sur de Tamaulipas. Tesis de Maestría, Facultad
Tampico region, México. Coastal Studies Institute, Loui-
González-Medrano, F. 1972. La vegetación del nordeste de
Puig, H. 1976. Vegetation de la Huasteca, Mexique. Mission
Tamaulipas. Anal. Inst. Biol. Mex. Ser. Bot. 43: 11-50.
Archeologique et Ethnologique Francaise du Mexique.
Griffin, E.A., Hopkins, A.J.M. & Hnatiuk, R.J. 1983. Re-
Coll. Etud. Mesoam. 5: 214-222.
gional variation in mediterranean-type shrublands near
Rejmanek, M. 1989. Invasibility of plant communities. In:
Eneabba, south-eastern Australia. Vegetatio 52: 103-127.
Drake, J.A., H.A. Mooney,H.A., di Castri, F., Groves,
Grootjans, A.P., Hartog, P.S., Fresco, L.F.M. & Esselink, H.
R.H., Kruger, F.J., Rejmanek, M. & Williamson, M. (eds.)
1991. Succession and fluctuation in a wet dune slack in
Biological Invasions. A global perspective. pp. 369-388 J.
relation to hydrological changes. J. Veg. Sci. 2: 545-554.
Guevara, S.S. 1982. Ecología de la vegetación de dunas
Romano, G.B. 1990. Invasibility of a mixed hardwood forest
costeras: esquema de investigación. Biotica 7: 603-610.
by Eupatorium capillifolium and E. compositifolium. M.Sc.
Hengeveld, R. 1989. Dynamics of biological invasions.
Sauer, J. D. 1967. Geographic reconnaissance of seashore
Huston, M.A. 1994. Biological diversity. The coexistence ofvegetation along the Mexican Gulf Coast. Coastal Studies
species on changing landscapes. Cambridge University
Institute, Louisiana State Univ. Techn. Rep. 56.
Sykes, M.T. & Wilson, B.J. 1987. The vegetation of a New
Johnson, A.F. & Barbour, M.G. 1990. Dunes and maritime
Zealand dune slack. Vegetatio 71: 13-19.
forests. In: Myers, R.L. & Ewel, J.J. (ed.) Ecosystems of
van der Laan, D. 1979. Spatial and temporal variation in the
Florida, pp. 429-480. Univ. of Central Florida Press,
vegetation of dune slacks in relation to the ground water
Kruger, F.J. 1977. Invasive woody plants in the Cape fynbos
van Wilgen, B.W. & Richardson, D.M. 1985. The effects of
with special reference to the biology and control of Pinus
alien shrub invasions on vegetation structure and fire
pinaster. Proceedings of the Second National Weeds Con-
behaviour in South African fynbos shrublands: a simula-
ference of South Africa. A.A. Balkema, Cape Town.
tion study. J. Appl. Ecol. 22: 955-966.
MacDonald, I.A.W. & Frame, G.W. 1988. The invasion of
introduced species into nature reserves in tropical savannasand dry woodlands. Biol. Conserv. 44: 67-93.
Mack, R.N. 1985. Invading plants: their potential contribution
to population biology. In: White, J (ed.) Studies in PlantDemography: a Festschrift for John Harper, pp. 127-141. Academic Press, London.
Maun, M.A. & Lapierre, J. 1984. The effects of burial by sand
on Ammophila arenaria. J. Ecol. 72: 827-829.
CHAMPAGNE & SPARKLING £GLASS £BOTTLE £BOTTLE Lanson Black Label, France A young, lively style with a lingering lemon character with slight biscuity aromas. . . . . . . . . . . . . . . . . . . . . . . . . . . 8.50 14.00 49.00 Lanson White Label, France Aromas of white flowers and summer pear on the nose. Soft & delicate on the palate, flavours of white fruit develop and c
T h e New E n g l a n d Jo u r n a l o f Me d i c i n ebefore pregnancy. Furthermore, for pregnant wom-en with certain conditions once believed to be in-compatible with pregnancy, such as systemic lupusA L A S T A I R J . J . W O O D , M . D. , Editor erythematosus and heart diseases, the outcome ofpregnancy has improved dramatically in the past fewdecades.4 DRUGS IN PREGNANCY In this arti
 Castillo, S.A. & Moreno-Casasola, P.
of 10-20% for the sites) which can have low or high
nity invasibility varies according to the characteristics
population numbers on a site and are frequently charac-
of the community and the life history traits of the po-
teristic of other community types. Huston (1994) con-
tential invaders (Ewel 1986). Disturbed communities
siders that invasions potentially lead to an increase in
are more often subject to invasions than intact commu-
species richness, as the invading species are added to the
nities and disturbance regime (type, frequency, dura-
existing species pool, although numerous examples dem-
tion and magnitude of disturbance) is recognized as the
onstrate that they can also lead to extinctions.
Castillo, S.A. & Moreno-Casasola, P.
of 10-20% for the sites) which can have low or high
nity invasibility varies according to the characteristics
population numbers on a site and are frequently charac-
of the community and the life history traits of the po-
teristic of other community types. Huston (1994) con-
tential invaders (Ewel 1986). Disturbed communities
siders that invasions potentially lead to an increase in
are more often subject to invasions than intact commu-
species richness, as the invading species are added to the
nities and disturbance regime (type, frequency, dura-
existing species pool, although numerous examples dem-
tion and magnitude of disturbance) is recognized as the
onstrate that they can also lead to extinctions.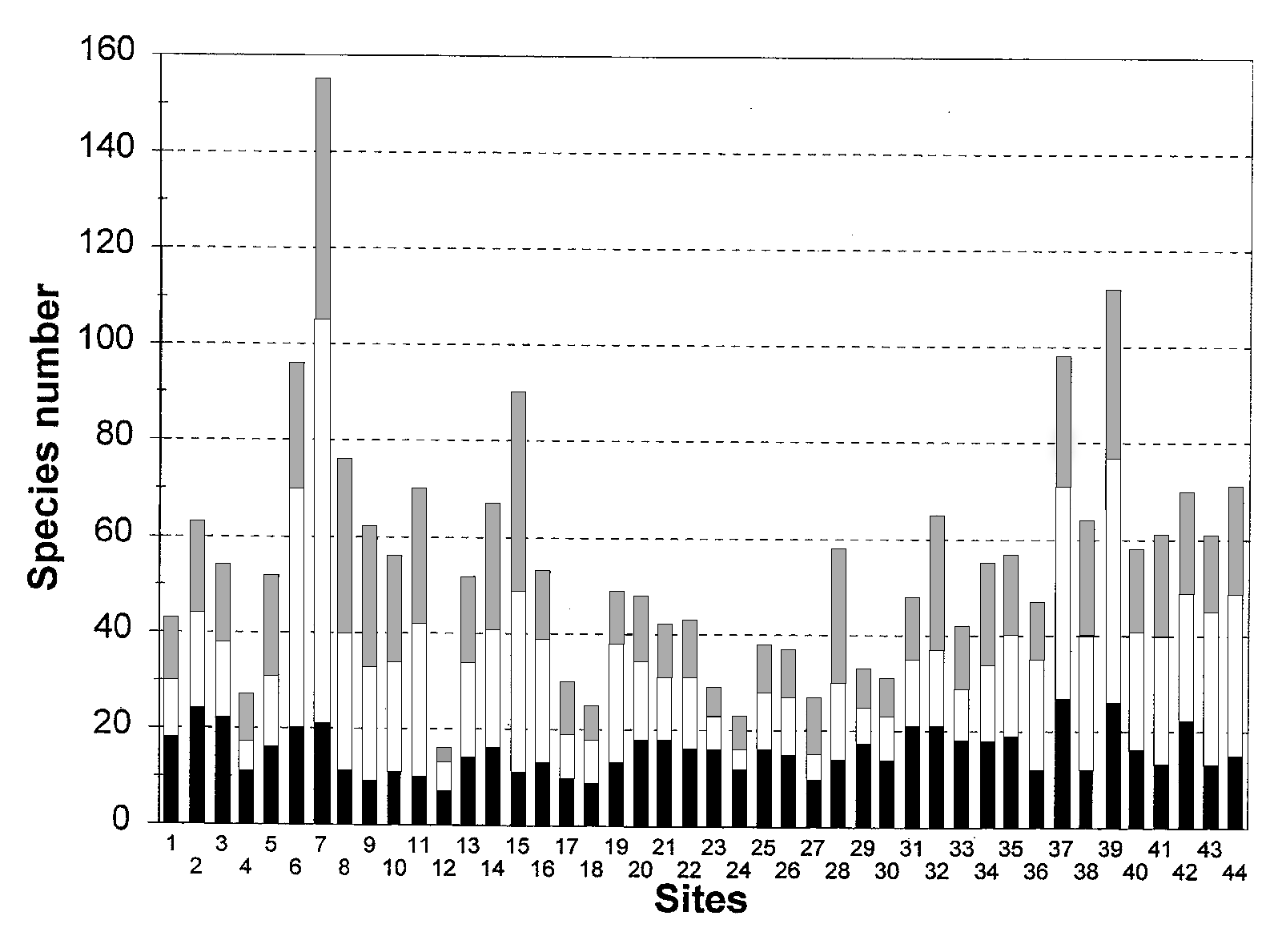 Castillo, S.A. & Moreno-Casasola, P.
Fig. 2. Species richness along the 44 Gulf and Caribbean sites. Black low parts of columns represent coastal species (C), open middle
Castillo, S.A. & Moreno-Casasola, P.
Fig. 2. Species richness along the 44 Gulf and Caribbean sites. Black low parts of columns represent coastal species (C), open middle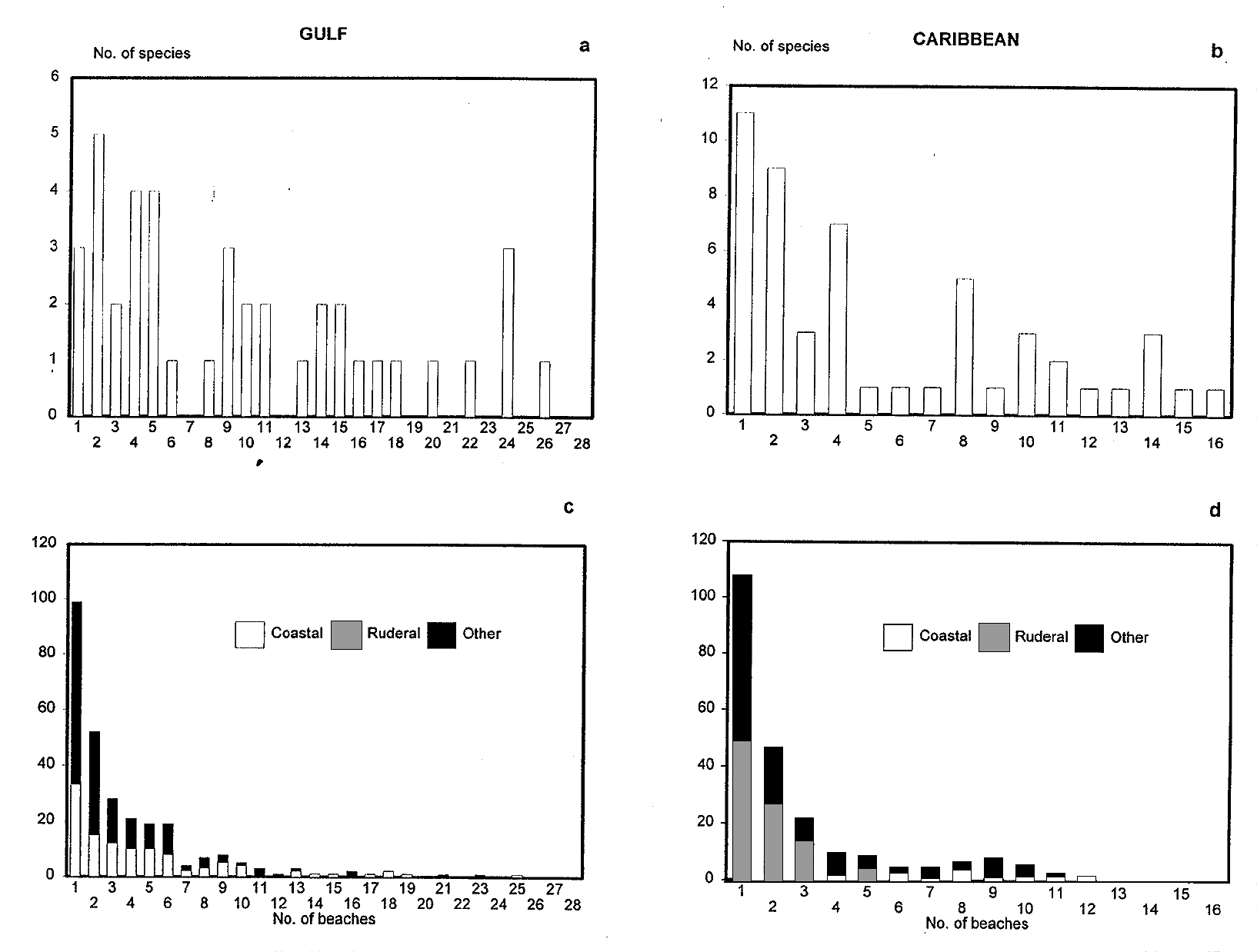 - Coastal sand dune vegetation: an extreme case of species invasion -
Fig. 3. Frequency distribution of coastal species (C) along 44 sites on the Gulf (a) and Caribbean (b). Frequency distribution of
- Coastal sand dune vegetation: an extreme case of species invasion -
Fig. 3. Frequency distribution of coastal species (C) along 44 sites on the Gulf (a) and Caribbean (b). Frequency distribution of