Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
People.biology.ucsd.edu
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, July 1998, p. 1778–1782
Copyright 1998, American Society for Microbiology. All Rights Reserved.
NorM, a Putative Multidrug Efflux Protein, of Vibrio parahaemolyticus
and Its Homolog in Escherichia coli
YUJI MORITA,1 KAZUYO KODAMA,1 SUMIKO SHIOTA,1 TOMOYUKI MINE,1 ATSUKO KATAOKA,1
TOHRU MIZUSHIMA,1 AND TOMOFUSA TSUCHIYA1,2,*
Department of Microbiology, Faculty of Pharmaceutical Sciences,1 and Gene Research Center,2
Okayama University, Tsushima, Okayama 700-8530, Japan
Received 17 February 1998/Returned for modification 27 March 1998/Accepted 6 May 1998
We found that cells of Vibrio parahaemolyticus possess an energy-dependent efflux system for norfloxacin. We cloned a gene for a putative norfloxacin efflux protein from the chromosomal DNA of V. parahaemolyticus by using an Escherichia coli mutant lacking the major multidrug efflux system AcrAB as the host and sequenced the gene (norM). Cells of E. coli transformed with a plasmid carrying the norM gene showed elevated energy- dependent efflux of norfloxacin. The transformants showed elevated resistance not only to norfloxacin and ciprofloxacin but also to the structurally unrelated compounds ethidium, kanamycin, and streptomycin. These results suggest that this is a multidrug efflux system. The hydropathy pattern of the deduced amino acid se- quence of NorM suggested the presence of 12 transmembrane domains. The deduced primary structure of NorM showed 57% identity and 88% similarity with that of a hypothetical E. coli membrane protein, YdhE. No reported drug efflux protein in the sequence databases showed significant sequence similarity with NorM. Thus, NorM seems to be a novel type of multidrug efflux protein. We cloned the ydhE gene from E. coli. Cells of E. coli transformed with the cloned ydhE gene showed elevated resistance to norfloxacin, ciprofloxacin, acriflavine, and tetraphenylphosphonium ion, but not to ethidium, when MICs were measured. Thus, it seems that NorM and YdhE differ somehow in substrate specificity.
Drug efflux from cells is one of the major mechanisms of
(18). Energy metabolism and energy coupling in membranes of
drug resistance in bacteria. Many drug efflux systems are
this microorganism are unique (11). Cells of V. parahaemolyti-
known in many bacteria (4, 22, 31). Four major groups of
cus show some natural resistance to some antimicrobial agents.
drug extrusion systems are known (4, 22, 31), i.e., the MF
Thus, we were interested in drug efflux systems of V. parahae-
(major facilitator) family, the SMR (small multidrug resis-
molyticus. Cells of V. parahaemolyticus may possess unique
tance) family, the RND (resistance nodulation cell division)
drug efflux systems. During the course of our studies, we found
family, and the ABC (ATP binding cassette) family. Mem-
that cells of V. parahaemolyticus possess an energy-dependent
brane transporters of the MF family possess 12 to 14 trans-
efflux system for norfloxacin, a widely used new quinolone an-
membrane domains. For example, Bcr (Escherichia coli) (1),
timicrobial agent. Here we report the cloning and sequencing
EmrB (E. coli) (14), EmrD (E. coli) (20), NorA (Staphylococ-
of the gene for and characterization of a putative norfloxacin
cus aureus) (34), QacA (S. aureus) (26), and Bmr (Bacillus
efflux protein of V. parahaemolyticus and the cloning and char-
subtilis) (21) are members of this family, and these systems
acterization of the homolog gene found in E. coli.
mediate drug extrusion with different specificities. Transport-
ers of the SMR family are rather small and usually possess four
MATERIALS AND METHODS
transmembrane domains. Smr (or QacC) (S. aureus) (8, 13),
QacE (Klebsiella aerogenes) (23), and EmrE (E. coli) (33) be-
Bacteria and growth. V. parahaemolyticus AQ3334 (30) and E. coli TG1 [⌬(lac-
long to this family. An electrochemical potential of Hϩ across
pro) supE thi hsd⌬5/FЈtraD36 proAϩBϩ lacIq lacZ⌬M15], a derivative of K-12,
and KAM2 and KAM3, derivatives of TG1, were used in this study. Cells of
cell membranes seems to be the driving force for drug efflux
V. parahaemolyticus AQ3334 were grown in LB medium (17) under aerobic
by the MF and SMR family transporters. Transporters of the
conditions at 37°C, and E. coli cells were grown in L medium (12) under aerobic
RND family consist of several subunits (usually three), and an
conditions at 37°C. Where indicated, drugs were added to the medium. Cell
outer membrane protein(s) is involved in the drug transport.
growth was monitored turbidimetrically at 650 nm. Isolation of mutants. KAM3, which has a deletion in the chromosomal acrAB
AcrAB (E. coli) (16) and MexAB (Pseudomonas aeruginosa)
genes, was obtained as follows. We tried to disrupt the acrAB genes, which code
(24) are examples of members of this family. Energy coupling
for the major multidrug efflux system of E. coli, by inserting the Mu phage. Cells
in this family is not very clear. However, it is clear that at least
of E. coli TG1 were infected with the Mud(Apr lac)I phage (5, 29). Cells were
an electrochemical potential of Hϩ across cell membranes is
diluted with L broth and spread onto an agar plate containing L broth and
40-g/ml ampicillin. After incubation at 37°C for 12 h, colonies were picked up
involved in driving the drug efflux (16). Transporters of the
and replica plated by using (i) an agar plate containing L broth and 40-g/ml
ABC family utilize ATP as the energy source. LmrA (Lacto-
ampicillin and (ii) an agar plate containing L broth, 40-g/ml ampicillin, and
coccus lactis) (32) and MsrA (S. aureus) (25) are members of
50-g/ml methylene blue, an antimicrobial basic dye and substrate for the AcrAB
system (16). We obtained two colonies which grew in the presence of methylene
blue. One of them was designated KAM2. Thereafter, the Mu phage region was
Vibrio parahaemolyticus, a slightly halophilic marine bacte-
removed from the chromosome of KAM2 cells by heat induction at 42°C (5, 29),
rium, is one of the major causes of food poisoning in Japan
and colonies that were sensitive to both ampicillin and methylene blue were
obtained. One of the colonies was designated KAM3. The KAM3 cells, as
well as the KAM2 cells, were sensitive to many drugs that are known as sub-
strates of the AcrAB system, although TG1 cells were resistant. The deletion
* Corresponding author. Mailing address: Department of Microbi-
in the acrAB region in KAM3 was confirmed by Southern blot analysis (data
ology, Faculty of Pharmaceutical Sciences, Okayama University, Tsu-
shima, Okayama 700-8530, Japan. Phone and Fax: 81-86-251-7957. Assay of norfloxacin accumulation in cells. V. parahaemolyticus cells were
E-mail: tsuchiya@pharm.okayama-u.ac.jp.
grown in the LB broth supplemented with 40 mM potassium lactate. The cells
DRUG EFFLUX SYSTEM IN V. PARAHAEMOLYTICUS
were harvested at the exponential phase of growth, washed with 0.2 M MOPS-
Tris buffer (pH 7.0) containing 10 mM MgSO4, and suspended in the same buffer
to 50 mg (wet weight)/ml. The assay mixture contained cells (10 mg [wet weight]/
ml) in the same buffer and 10 mM potassium lactate. After incubation at 25°C for
5 min, norfloxacin (100 M, final concentration) was added to initiate the assay.
Samples (1 ml each) were taken at intervals, centrifuged at 7,000 ϫ g for 30 s at
4°C, and washed once with the same buffer. Where indicated, carbonyl cyanide
m-chlorophenylhydrazone (CCCP) was added to the assay mixture at 100 M.
The pellet was suspended in 1 ml of 100 mM glycine-HCl (pH 3.0). The suspen-
sion was shaken vigorously for 1 h at room temperature and then centrifuged at
7,000 ϫ g for 5 min at room temperature. The supernatant was diluted twofold
with the same buffer, and the fluorescence was measured with excitation at 277
nm and emission at 448 nm (10, 19) with a Hitachi F2000 fluorescence spectro-
Assay of ethidium accumulation in cells. Cells were prepared and suspended
as described above for the norfloxacin accumulation assay. Potassium lactate
(final concentration 20 mM) was added to the cell suspension (0.5 mg of protein/
ml), and the cell suspension was kept at 25°C for 5 min with gentle stirring.
Ethidium bromide was added at 10 M to the cell suspension to initiate the
assay. CCCP was added at 100 M where indicated. The fluorescence of the
assay mixture was measured with excitation and emission wavelengths of 500 and
Gene cloning and sequencing. The gene responsible for norfloxacin efflux was
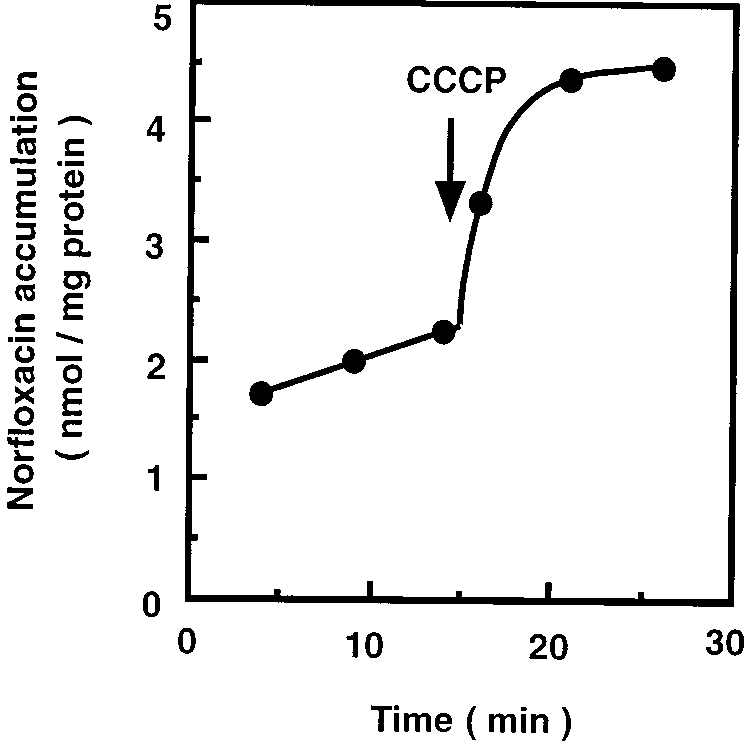
FIG. 1. Accumulation of norfloxacin in cells of V. parahaemolyticus. V. para-
cloned from V. parahaemolyticus cells as follows. Chromosomal DNA was pre-
haemolyticus cells were grown in LB medium supplemented with 40 mM potas-
pared from cells of V. parahaemolyticus by the method of Berns and Thomas (2).
sium lactate. Norfloxacin was added to the cell suspensions at a final concentra-
The DNA was partially digested with Sau3AI, and fragments of 4 to 10 kbp were
tion of 100 M. After 15 min, CCCP was added to the suspensions at a final
separated by sucrose density gradient centrifugation. The DNA fragments were
concentration of 100 M. Portions were removed at the times shown, and the
ligated into pBR322 (which had been digested with BamHI and dephosphory-
concentration of norfloxacin extracted from the cells was determined with a
lated with bacterial alkaline phosphatase) by using T4 DNA ligase. Competent
cells of E. coli KAM3 were transformed with the ligated hybrid plasmids and
were spread on agar plates containing L broth, 0.05-g/ml norfloxacin, 60-g/ml
ampicillin, and 1.5% agar. The plates were incubated at 37°C for 24 h, and the
RESULTS AND DISCUSSION
clones formed were picked up. Plasmids contained in the transformants were
isolated, reintroduced into KAM3 cells, and spread on the plates again. The
Norfloxacin efflux in cells of V. parahaemolyticus. We mea-
plates were incubated at 37°C for 24 h. Plasmids contained in the retransfor-
sured norfloxacin accumulation and tested the effect of an Hϩ
mants were prepared. One of the resulting hybrid plasmids that carried a gene
for the norfloxacin efflux system was designated pMVP3.
conductor, CCCP, on the accumulation in cells of V. parahae-
The DNA insert in pMVP3 was digested with several restriction endonucleases
molyticus. A certain level of norfloxacin accumulation in the
and subcloned into pBR322. The resulting hybrid plasmids were introduced into
cells was observed, and the accumulation level increased after
KAM3 cells, and the transformants were tested for sensitivity or resistance to
addition of CCCP (Fig. 1), suggesting the existence of a nor-
floxacin efflux system in V. parahaemolyticus that is driven by an
The nucleotide sequence was determined by the dideoxy chain termination
electrochemical potential of Hϩ. It is not clear whether Hϩ is
method (27) with a DNA sequencer (ALF Express, Pharmacia Biotech).
The ydhE gene of E. coli was cloned as follows. Chromosomal DNA was
directly or indirectly coupled to the drug efflux.
prepared from cells of E. coli KAM3 by the method of Berns and Thomas (2). Cloning of the gene for a putative norfloxacin efflux protein.
The DNA was digested with SspI and SphI, which should cut out the whole ydhE
We cloned a fragment of the chromosomal DNA of V. para-
gene (EMBL nucleotide sequence database accession no. AE000261), judging
haemolyticus, which enabled norfloxacin-hypersensitive E. coli
from the DNA sequence of the E. coli genome. The SspI-SphI fragments were
KAM3 cells to grow in the presence of norfloxacin. We ob-
ligated into pBR322 (which had been digested with EcoRV and SphI and de-
phosphorylated with bacterial alkaline phosphatase) by using T4 DNA ligase.
tained about 40 candidate hybrid plasmids. However, it seemed
Competent cells of E. coli KAM3, which are very sensitive to norfloxacin, were
that the DNA inserts in all of the candidate plasmids contained
transformed with the ligated hybrid plasmids and spread on plates containing L
the same DNA portion, judging from restriction maps of those
broth, 0.05-g/ml norfloxacin, 60-g/ml ampicillin, and 1.5% agar. The plates
plasmids. We tested whether the plasmids carry a gene(s) re-
were incubated at 37°C for 24 h, and the clones formed were picked up. Plasmids
sponsible for norfloxacin efflux or not by measuring norfloxa-
contained in the transformants were checked for whether they contained the
cin accumulation and the effect of CCCP in E. coli cells trans-
expected SspI-SphI fragment. One of the resulting hybrid plasmids that carried
the ydhE gene was designated pMEC2.
formed with the plasmids. A considerable level of norfloxacin
Sequence data were analyzed with GENETYX sequence analysis software
accumulation was observed with the host cells (E. coli KAM3).
(Software Development Co.). The SwissProt and GenBank databases were
Addition of CCCP increased the accumulation level to some
extent, supporting the presence of some norfloxacin efflux
Southern blot analysis. Chromosomal DNAs were prepare from cells of
system(s) in E. coli KAM3 (6). Cells harboring the plasmids
E. coli, V. parahaemolyticus, V. alginolyticus, P. aeruginosa, and S. aureus as
described above. The DNAs were digested with several restriction enzymes and
showed very low levels of norfloxacin accumulation compared
subjected to agarose gel electrophoresis. DNA fragments were blotted onto a
with those of the host cells. The norfloxacin accumulation in
Hybond-N (Amersham Co.) nylon membrane by the capillary blotting method as
cells harboring one of the plasmids, pMVP3, is shown in Fig. 2.
suggested by the manufacturer. The probes used were three DNA fragments
Addition of CCCP to the assay mixture greatly increased the
derived from the acrRAB genes of E. coli, which are shown below. Southern blot
accumulation level (Fig. 2). The norfloxacin accumulation level
analysis was performed with the enhanced-chemiluminescence detection system
after the addition of CCCP was very similar in the host cells
(Amersham Co.) as suggested by the manufacturer. Drugs susceptibility test. The MICs of drugs were determined in Mueller-
and the transformed cells. Thus, we conclude that the elevat-
Hinton broth (Difco) containing various drugs at various concentrations. Cells in
ed norfloxacin efflux is due to the gene(s) carried on plasmid
the test medium were incubated at 37°C for 24 h, and growth was judged
Inhibition of the drug efflux in KAM3/pMVP3 cells by
Other. Protein contents were determined by the method of Lowry et al. (15).
CCCP indicates that an electrochemical potential of Hϩ is the
The chemicals and enzymes used in this study were from commercial sources. Nucleotide sequence accession number. The nucleotide sequence data re-
driving force for the drug extrusion. Some drug/Hϩ antiport-
ported in this paper have been deposited in the DDBJ, EMBL, and GenBank
ers, such as the tetracycline/Hϩ antiporter (9), are known in
nucleotide sequence databases under accession no. AB010463.
microbial cells. It is very likely that the norfloxacin extrusion
FIG. 3. Restriction maps of pMVP3 and its deletion derivatives. DNA re-
gions derived from V. parahaemolyticus chromosomal DNA and carried by each
plasmid are shown. Growth of E. coli KAM3 cells harboring each plasmid on an
agar plate containing L broth, 0.05-g/ml norfloxacin, and 60-g/ml ampicillin is
shown on the right. Plus signs indicate that cells grew, and minus signs indicate
FIG. 2. Accumulation of norfloxacin in host and transformant cells. Cells of
that cells did not grow. The arrow shows the position and direction of the norME. coli KAM3 and KAM3/pMVP3 were grown in L medium supplemented with
40 mM potassium lactate. Norfloxacin was added to the cell suspensions at a final
concentration of 100 M. After 15 min, CCCP was added to the suspensions at
a final concentration of 100 M. Portions were removed at the times shown, and
the concentration of norfloxacin extracted from the cells was determined with a
fluorescence spectrophotometer. Symbols: E, KAM3; F, KAM3/pMVP3.
We checked whether the cloned DNA fragment is really
from V. parahaemolyticus, and whether a similar gene is pres-
ent in other bacteria, by Southern blot analysis. A DNA frag-
ment derived from pMVP36 was used as a probe. We detected
system in KAM3/pMVP3 is a drug/ion antiporter. However, it
a dense hybridized band with chromosomal DNA of V. para-
is not clear whether this system utilizes Hϩ as the counterion
haemolyticus (data not shown). The position of the band was
for the antiport. Many ion-coupled systems in membranes of
exactly the same as that of the control (pMVP36). Thus, we
vibrios utilize Naϩ instead of Hϩ (28). Although cells of
believe that the DNA insert in pMVP36 is from the chromo-
V. parahaemolyticus possess a primary respiratory Naϩ pump
somal DNA of V. parahaemolyticus. We also detected a less
(30), Naϩ-coupled membrane processes, such as Naϩ/solute
dense band with chromosomal DNA of V. alginolyticus. Thus,
symport, are sensitive to an Hϩ conductor (unpublished re-
it seems that cells of V. alginolyticus possess a similar gene.
sults). Therefore, we cannot distinguish between an Hϩ-cou-
Chromosomal DNAs from other bacteria tested, P. aeruginosa
pled system and an Naϩ-coupled system from the effects of an
Hϩ conductor. We tested the effect of Naϩ on the activity of
norfloxacin extrusion in cells of both V. parahaemolyticus and
E. coli transformed with pMVP3. However, no clear effect was
TABLE 1. Susceptibilities of study strains to different compounds
observed (data not shown). We also tried to detect norfloxacin/
Hϩ antiport by the quinacrine fluorescence quenching method
with everted membrane vesicles prepared from cells of KAM3/
pMVP3. However, we were unable to detect Hϩ efflux due to
norfloxacin influx (data not shown).
Several plasmids carrying different portions of pMVP3 were
constructed, and the ability to confer norfloxacin resistance on
E. coli KAM3 was tested (Fig. 3). Among the plasmids that con-
ferred norfloxacin resistance, plasmid pMVP36 possessed the
shortest DNA insert derived from V. parahaemolyticus DNA
We tested the susceptibility of cells of KAM3/pMVP36 to
many drugs (Table 1). pMVP36 made KAM3 cells resistant to
rather hydrophilic new quinolones, such as norfloxacin and
ciprofloxacin, but not to hydrophobic quinolones, such as spa-
floxacin and nalidixic acid. Cells harboring pMVP36 were also
resistant to structurally different antimicrobial agents, such as
ethidium and streptomycin (and kanamycin). Thus, it seems
that pMVP36 carries a gene for multidrug resistance. Our re-
sults are consistent with the idea that the gene encodes a pro-
tein mediating the extrusion of norfloxacin and ethidium from
cells. In fact, we observed elevated ethidium efflux in cells of
KAM3/pMVP36 compared with cells of KAM3 (data not shown).
DRUG EFFLUX SYSTEM IN V. PARAHAEMOLYTICUS
FIG. 4. Amino acid sequence alignment. The deduced amino acid sequence of V. parahaemolyticus NorM (NorM VPARA) and the amino acid sequences of E. coli
YdhE (YdhE ECOLI) and H. influenzae YdhE (YdhE HAEIN) are aligned. The numbers on both sides refer to the position of the nearest residue on each line.
Asterisks and dots indicate residues in YdhE of E. coli that are identical and similar to those in NorM, respectively.
and S. aureus, showed no hybridized band under our experi-
residues (Fig. 4) with a molecular mass of 49,422 Da and is very
rich in hydrophobic residues, indicating that the protein is a
Gene and protein sequences. We determined the sequence
membrane protein. A hydropathy analysis done by the method
of 2,243 nucleotides of the DNA insert in pMVP36. We found
of Eisenberg et al. (7) revealed that NorM possesses 12 hydro-
only one open reading frame, which was preceded by a Shine-
phobic regions which may be transmembrane domains (data
Dalgarno sequence in this region, which is likely to be the gene
not shown). Therefore, it is likely that NorM is a member of
(which we designated norM) encoding the norfloxacin efflux
protein (NorM). Several promoterlike sequences were present
Homolog of NorM. We searched for amino acid sequence
upstream of norM. The amino acid sequence deduced from the
homology between NorM and the reported sequences in a pro-
norM gene revealed that NorM consists of 456 amino acid
tein sequence database (SwissProt). No sequence homology
or similarity was observed between NorM and the reported
iya. 1994. Properties of the Naϩ/Hϩ antiporter in Vibrio parahaemolyticus.
members of the MF family. The first extrusion system for
J. Biochem. 115:1162–1165.
norfloxacin to be reported in microbial cells is the NorA system
12. Lennox, E. S. 1955. Transduction of linked genetic characters of host by
of S. aureus (34). We found no sequence similarity between
bacteriophage P1. Virology 1:190–206.
13. Littlejohn, T. G., D. DiBerardino, L. J. Messerotti, S. J. Spiers, and R. A.
NorM and NorA. However, two hypothetical proteins, YdhE
Skurray. 1991. Structure and evolution of a family of genes encoding anti-
of E. coli and that of Haemophilus influenzae, showed high
septic and disinfectant resistance in Staphylococcus aureus. Gene 101:59–66.
sequence similarity with NorM. The sequence identity and
14. Lomovskaya, O., and K. Lewis. 1992. Emr, an Escherichia coli locus for
similarity between NorM and YdhE of E. coli were 57 and
multidrug resistance. Proc. Natl. Acad. Sci. USA 89:8938–8942.
15. Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein
88%, respectively, and those between NorM and YdhE of H.
measurement with the Folin phenol reagent. J. Biol. Chem. 193:265–275. influenzae were 43 and 85%, respectively. Figure 4 shows an
16. Ma, D., D. N. Cook, M. Alberti, N. G. Pon, H. Nikaido, and J. E. Hearst.
alignment of the deduced amino acid sequences of NorM and
1993. Molecular cloning and characterization of acrA and acrE genes of
these YdhE proteins. Judging from the high sequence identity
Escherichia coli. J. Bacteriol. 175:6299–6313.
17. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: a
and similarity, it seems that NorM and these two YdhE pro-
laboratory manual, p. 68. Cold Spring Harbor Laboratory, Cold Spring
We cloned the ydhE gene from the chromosomal DNA of
18. Miwatani, T., and Y. Takeda. 1976. Food poisoning due to Vibrio parahae- E. coli KAM3. Cells of E. coli KAM3 harboring a hybrid plas-
molyticus in Japan, p. 22–25. In T. Miwatani and Y. Takeda (ed.), Vibrio
mid carrying the ydhE gene, pMEC2, showed elevated resis-
parahaemolyticus, a causative bacterium of food poisoning. Saikon Publish-
tance to several drugs, such as norfloxacin, ciprofloxacin, and
19. Mortimer, P. G., and L. J. Piddock. 1991. A comparison of methods used for
acriflavine, and the tetraphenylphosphonium ion, judging from
measuring the accumulation of quinolones by Enterobacteriaceae, Pseudomo-
the MICs (Table 1). Some increase in the MICs of kanamycin
nas aeruginosa and Staphylococcus aureus. J. Antimicrob. Chemother. 28:
and streptomycin was observed. Surprisingly, no MIC increase
20. Naroditskaya, V., M. J. Schlosser, N. Y. Fang, and K. Lewis. 1993. An E. coli
was observed with ethidium, which is a good substrate for the
gene emrD is involved in adaptation to low energy shock. Biochem. Biophys.
Res. Commun. 196:803–809.
Another hypothetical protein (YojI) of B. subtilis (EMBL
21. Neyfakh, A. A., V. E. Bidnenko, and L. B. Chen. 1991. Efflux-mediated
database accession no. Z99114) had an amino acid sequence
multidrug resistance in Bacillus subtilis: similarities and dissimilarities with
the mammalian system. Proc. Natl. Acad. Sci. USA
that was similar to that of NorM (35% identity and 77% sim-
88:4781–4785.
22. Nikaido, H. 1996. Multidrug efflux pumps of gram-negative bacteria. J.
Bacteriol. 178:5853–5859.
23. Paulsen, I. T., T. G. Littlejohn, P. Rådstro¨m, L. Sundstro¨m, O. Sko¨ld, G. Swedberg, and R. A. Skurray. 1993. The 3Ј conserved segment of integrons ACKNOWLEDGMENTS
contains a gene associated with multidrug resistance to antiseptics and dis-
infectants. Antimicrob. Agents Chemother. 37:761–768.
This research was supported by a grant from the Ministry of Edu-
24. Poole, K., D. E. Heinrichs, and S. Neshat. 1993. Cloning and sequence
cation, Science and Culture of Japan.
analysis of an EnvCD homologue in Pseudomonas aeruginosa: regulation by
iron and possible involvement in the secretion of the siderophore pyover-
REFERENCES
dine. Mol. Microbiol. 10:529–544. Bentley, J., L. S. Hyatt, K. Ainley, J. H. Parish, R. B. Herbert, and G. R. Ross, J. I., E. A. Eady, J. H. Cove, W. J. Cunliffe, S. Baumberg, and J. C. White. 1993. Cloning and sequence analysis of an Escherichia coli gene Wootton. 1990. Inducible erythromycin resistance in staphylococci is en-
coded by a member of the ATP-binding transport super-gene family. Mol. 127:117–120.
2. Berns, K. I., and C. A. J. Thomas. 1965. Isolation of high molecular weight
Microbiol. 4:1207–1214.
DNA from Haemophilus influenzae. J. Mol. Biol. 11:476–490.
26. Rouch, D. A., D. S. Cram, D. DiBerardino, T. G. Littlejohn, and R. A.
3. Bolhuis, H., D. Molenaar, G. Poelarends, H. W. van Veen, B. Poolman, Skurray. 1990. Efflux-mediated antiseptic resistance gene qacA from Staph- A. J. M. Driessen, and W. N. Konings. 1994. Proton motive force-driven and ylococcus aureus: common ancestry with tetracycline- and sugar-transport
ATP-dependent drug extrusion systems in multidrug-resistant Lactococcus
proteins. Mol. Microbiol. 4:2051–2062. lactis. J. Bacteriol. 176:6957–6964.
27. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencing with
4. Bolhuis, H., H. W. van Veen, B. Poolman, A. J. Driessen, and W. N. Konings.
chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467.
1997. Mechanisms of multidrug transporters. FEMS Microbiol. Rev. 21:55–
28. Skulachev, V. P. 1989. The sodium cycle: a novel type of bacterial energetics.
J. Bioenerg. Biomembr. 21:635–647.
5. Casadaban, M. J., and S. N. Cohen. 1979. Lactose genes fused to exogenous
29. Tate, C. G., J. A. Muiry, and P. J. Henderson. 1992. Mapping, cloning,
promoters in one step using a Mu-lac bacteriophage: in vivo probe for
expression, and sequencing of the rhaT gene, which encodes a novel L-
transcriptional control sequences. Proc. Natl. Acad. Sci. USA 76:4530–4533.
rhamnose-Hϩ transport protein in Salmonella typhimurium and Escherichia
6. Cohen, S. P., D. C. Hooper, J. S. Wolfson, K. S. Souza, L. M. McMurry, and coli. J. Biol. Chem. 267:6923–6932. S. B. Levy. 1988. Endogenous active efflux of norfloxacin in susceptible
30. Tsuchiya, T., and S. Shinoda. 1985. Respiration-driven Naϩ pump and Naϩ Escherichia coli. Antimicrob. Agents Chemother. 32:1187–1191.
circulation in Vibrio parahaemolyticus. J. Bacteriol. 162:794–798.
7. Eisenberg, D., E. Schwarz, M. Komaromy, and R. Wall. 1984. Analysis of
31. van Veen, H. W., and W. N. Konings. 1997. Drug efflux proteins in multidrug
membrane and surface protein sequences with the hydrophobic moment
resistant bacteria. Biol. Chem. 378:769–777.
plot. J. Mol. Biol. 179:125–142.
32. van Veen, H. W., K. Venema, H. Bolhuis, I. Oussenko, J. Kok, B. Poolman,
8. Grinius, L., G. Dreguniene, E. B. Goldberg, C. H. Liao, and S. J. Projan. A. J. Driessen, and W. N. Konings. 1996. Multidrug resistance mediated by
1992. A staphylococcal multidrug resistance gene product is a member of a
a bacterial homolog of the human multidrug transporter MDR1. Proc. Natl.
new protein family. Plasmid 27:119–129.
Acad. Sci. USA 93:10668–10672.
9. Kaneko, M., A. Yamaguchi, and T. Sawai. 1985. Energetics of tetracycline
33. Yerushalmi, H., M. Lebendiker, and S. Schuldiner. 1995. EmrE, an Esche-
efflux system encoded by Tn10 in Escherichia coli. FEBS Lett. 193:194–198. richia coli 12-kDa multidrug transporter, exchanges toxic cations and Hϩ and
10. Kotera, Y., M. Watanabe, S. Yoshida, M. Inoue, and S. Mitsuhashi. 1991.
is soluble in organic solvents. J. Biol. Chem. 270:6856–6863.
Factors influencing the uptake of norfloxacin by Escherichia coli. J. Antimi-
34. Yoshida, H., M. Bogaki, S. Nakamura, K. Ubukata, and M. Konno. 1990.
crob. Chemother. 27:733–739.
Nucleotide sequence and characterization of the Staphylococcus aureus norA
11. Kuroda, T., T. Shimamoto, K. Inaba, T. Kayahara, M. Tsuda, and T. Tsuch-
gene, which confers resistance to quinolones. J. Bacteriol. 172:6942–6949.
(Código do Mercado de Valores Mobiliários) Compete ao Banco de Cabo Verde, através da Auditoria Geral do Mercado de Valores Mobiliários a fiscalização das ofertas públicas de aquisição ou de venda. (Interrupção da negociação em bolsa) 1. A Auditoria Geral do Mercado de Valores Mobiliários poderá interromper a negociação em bolsa dos valores mobiliários objecto de ofertas públ
GAGLIANO: MOVING LASIX DEBATE FROM 'HYPERBOLE AND FEAR- MONGERING' TO FACTS ABOUT COMPETITION by Paulick Report Staff | 01.28.2013 | 2:11pm The following letter to the editor was sent by Jim Gagliano, president and chief operating officer of the Jockey Club, in response to a Thoroughbred Daily News report on a panel earlier this month in California. The scientists invited to speak on the p
 DRUG EFFLUX SYSTEM IN V. PARAHAEMOLYTICUS
were harvested at the exponential phase of growth, washed with 0.2 M MOPS-
Tris buffer (pH 7.0) containing 10 mM MgSO4, and suspended in the same buffer
to 50 mg (wet weight)/ml. The assay mixture contained cells (10 mg [wet weight]/
ml) in the same buffer and 10 mM potassium lactate. After incubation at 25°C for
5 min, norfloxacin (100 M, final concentration) was added to initiate the assay.
DRUG EFFLUX SYSTEM IN V. PARAHAEMOLYTICUS
were harvested at the exponential phase of growth, washed with 0.2 M MOPS-
Tris buffer (pH 7.0) containing 10 mM MgSO4, and suspended in the same buffer
to 50 mg (wet weight)/ml. The assay mixture contained cells (10 mg [wet weight]/
ml) in the same buffer and 10 mM potassium lactate. After incubation at 25°C for
5 min, norfloxacin (100 M, final concentration) was added to initiate the assay.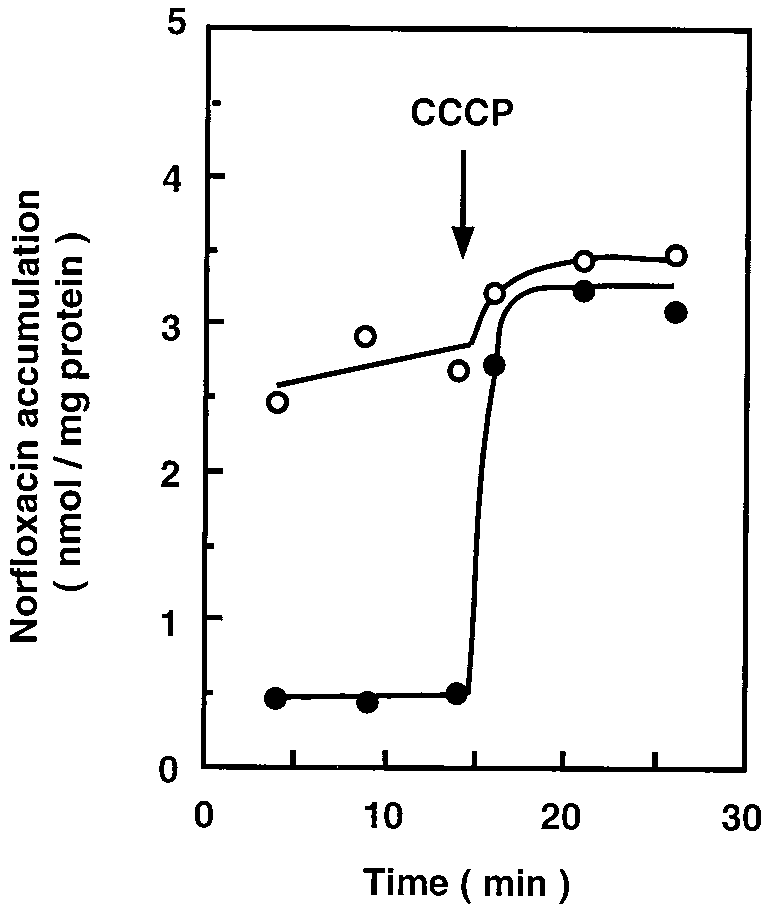
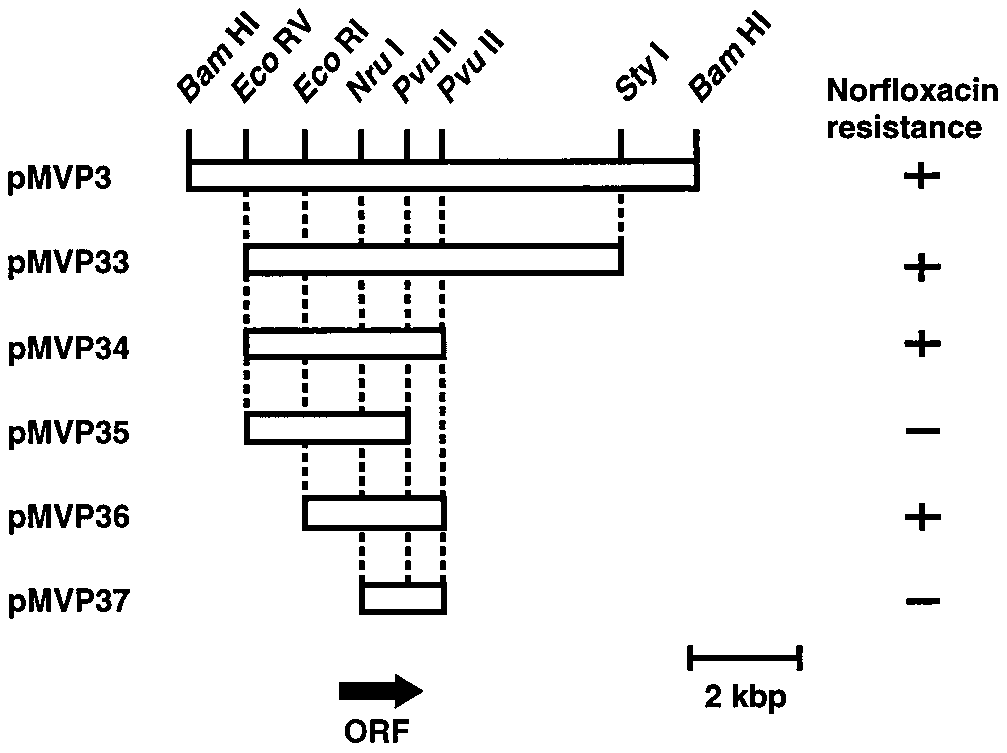 FIG. 3. Restriction maps of pMVP3 and its deletion derivatives. DNA re-
gions derived from V. parahaemolyticus chromosomal DNA and carried by each
plasmid are shown. Growth of E. coli KAM3 cells harboring each plasmid on an
agar plate containing L broth, 0.05-g/ml norfloxacin, and 60-g/ml ampicillin is
shown on the right. Plus signs indicate that cells grew, and minus signs indicate
FIG. 2. Accumulation of norfloxacin in host and transformant cells. Cells of
that cells did not grow. The arrow shows the position and direction of the norM
E. coli KAM3 and KAM3/pMVP3 were grown in L medium supplemented with
40 mM potassium lactate. Norfloxacin was added to the cell suspensions at a final
concentration of 100 M. After 15 min, CCCP was added to the suspensions at
a final concentration of 100 M. Portions were removed at the times shown, and
the concentration of norfloxacin extracted from the cells was determined with a
fluorescence spectrophotometer. Symbols: E, KAM3; F, KAM3/pMVP3.
FIG. 3. Restriction maps of pMVP3 and its deletion derivatives. DNA re-
gions derived from V. parahaemolyticus chromosomal DNA and carried by each
plasmid are shown. Growth of E. coli KAM3 cells harboring each plasmid on an
agar plate containing L broth, 0.05-g/ml norfloxacin, and 60-g/ml ampicillin is
shown on the right. Plus signs indicate that cells grew, and minus signs indicate
FIG. 2. Accumulation of norfloxacin in host and transformant cells. Cells of
that cells did not grow. The arrow shows the position and direction of the norM
E. coli KAM3 and KAM3/pMVP3 were grown in L medium supplemented with
40 mM potassium lactate. Norfloxacin was added to the cell suspensions at a final
concentration of 100 M. After 15 min, CCCP was added to the suspensions at
a final concentration of 100 M. Portions were removed at the times shown, and
the concentration of norfloxacin extracted from the cells was determined with a
fluorescence spectrophotometer. Symbols: E, KAM3; F, KAM3/pMVP3. DRUG EFFLUX SYSTEM IN V. PARAHAEMOLYTICUS
FIG. 4. Amino acid sequence alignment. The deduced amino acid sequence of V. parahaemolyticus NorM (NorM VPARA) and the amino acid sequences of E. coli
YdhE (YdhE ECOLI) and H. influenzae YdhE (YdhE HAEIN) are aligned. The numbers on both sides refer to the position of the nearest residue on each line.
DRUG EFFLUX SYSTEM IN V. PARAHAEMOLYTICUS
FIG. 4. Amino acid sequence alignment. The deduced amino acid sequence of V. parahaemolyticus NorM (NorM VPARA) and the amino acid sequences of E. coli
YdhE (YdhE ECOLI) and H. influenzae YdhE (YdhE HAEIN) are aligned. The numbers on both sides refer to the position of the nearest residue on each line.