Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Untitled
Protective effect of ginsenosides Rg2 and Rh1on oxidation-induced impairment oferythrocyte membrane properties
Keiichi Samukawa a, Yoji Suzuki b, Nobutaka Ohkubo b, Mamoru Aoto b,Masahiro Sakanaka a and Noriaki Mitsuda b,∗a Department of Functional Histology, Graduate School of Medicine, Ehime University, Toon,Ehime, Japanb Department of Physiology, Graduate School of Medicine, Ehime University, Toon, Ehime, JapanAbstract. The extract from Panax ginseng has been reported to improve the microcirculation in various organs. However, the mechanisms underlying this phenomenon are still poorly understood. In the present study, using the rheological properties of erythrocytes as an index, we have screened the components of Panax ginseng extract and identified Rg2 and Rh1 as the active ingredients. These two ginsenosides prevented the oxidative stress-induced elevation of erythrocyte suspension viscosity and the impairment of erythrocyte elongation in response to shear stress. Rg2 and Rh1 ginsenosides did not have antioxidant activity in an aqueous phase and did not inhibit the peroxidation of membrane lipids, either. However, they inhibited the oxidation-induced decrease of SH-groups in band 3 (anion exchanger-1), one of the important structural proteins of the erythrocyte membrane, but not in other structural proteins: bands 1 and 2 (spectrins), band 4.2 or band 5 (actin). These results suggest that ginsenosides Rg2 and Rh1 protect the rheological functions of erythrocytes against oxidative stress by preventing the oxidation of SH-groups in band 3 protein.
Keywords: Ginsenoside Rg2, ginsenoside Rh1, thiol group, band 3, oxidative stress, erythrocyte
1. Introduction
Circulating erythrocytes are constantly exposed to oxidative stress as a result of the high cellular
concentration of oxygen and hemoglobin, which is a source of the reactive oxygen species. Exposureto oxidative stress dramatically alters the function of the erythrocyte membrane, such as elevation ofviscosity, decrease of deformability, rouleaux formation and hemolysis [6,7,18]. In parallel with theseadverse functional changes, exposure to oxidative stress causes several types of molecular damage inerythrocytes, such as peroxidation of lipids, degradation of proteins, hemoglobin–spectrin cross-linking,glycosylation of proteins, inactivation of enzymes and perturbation of the membrane lipid bilayer.
The extract from the root of Panax ginseng C.A. Meyer has been used as traditional medicine in Asian
countries for more than 2000 years. The major active components, called ginsenosides, are derivatives ofthe dammarane-type triterpene, except for Ro (the only oleanolic acid-type ginsenoside). According to
*Address for correspondence: Noriaki Mitsuda, Department of Physiology, Graduate School of Medicine, Ehime University,
Shitsukawa, Toon, Ehime 791-0295, Japan. Tel.: +81 89 960 5245; Fax: +81 89 960 5246; E-mail: mitsuda@m.ehime-u.ac.jp.
0006-355X/08/$17.00 2008 – IOS Press and the authors. All rights reserved
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
the position of the sugar moieties, they are classified into 2 groups: the protopanaxadiol and protopanax-atriol groups. The protopanaxadiol group includes ginsenosides Rb1, Rb2, Rc, Rd, Rg3, Rh2 and Rh3 inwhich the sugar moieties are attached to the 3- the 20-position of the dammarane-type triterpene, whilethe protopanaxatriol group includes ginsenoside Re, Rf, Rg1, Rg2 and Rh1 in which the sugar moietiesare attached to the 6- and the 20-position of 3-OH-dammarane-type triterpene [2].
The extract from ginseng has often been reported to improve the microcirculation in various organs
[5,11,14,25,26] and also shown to have an antioxidant effect [12,13,15,19,32]. However, it is not welldescribed which of the ginsenosides are responsible for these beneficial effects of the ginseng extract. Among the ginsenosides, many researchers including us reported that ginsenoside Rb1 protects neuronal,endothelial and hepatic cells from oxidative stress [8,16,23,31]. So, we used ginsenoside Rb1 togetherwith vitamins C and E as a positive control of antioxidant.
In our previous study [27], we reported that exposure of erythrocytes to strong oxidative stress (gen-
erated by 2 mM FeSO4 with 10 mM ascorbate) led to activation of caspase-3 in the cells, which resultedin cleavage of band 3 protein and impairment of erythrocyte membrane functions, such as deformabilityand anion exchange. In that study, we also showed that inhibition of caspase-3 with the specific inhibitorsuppressed the digestion of band 3, and attenuated the functional damage. It should be noted, however,that the caspase-3 inhibitor did not completely inhibit the damage. These results suggested the existenceof mechanisms other than caspase-3, by which oxidative stress damages the functions of the erythrocytemembrane.
In the present study, we exposed the erythrocytes to a low-level oxidative stress (generated by 0.5 mM
FeSO4 with 5 mM ascorbate). Under such weak oxidative stress, band 3 protein was hardly cleaved butthe erythrocyte membrane functions were significantly impaired. Under these conditions, we investi-gated the mechanisms underlying the membrane damage, and whether the Panax ginseng componentsprotect the erythrocytes from oxidative stress-induced membrane dysfunction. Using erythrocyte sus-pension viscosity and erythrocyte deformability as indices of rheological properties, we identified Rg2and Rh1 as the anti-oxidative components of Panax ginseng extract. 2. Materials and methods
Ferrous sulfate (FeSO4), ascorbic acid, trichloroacetic acid (TCA), thiobarbituric acid (TBA) and
butylated hydroxytoluene (BHT) were purchased from Wako pure chemicals (Osaka, Japan), Dex-tran T-40 from Pharmacia (Uppsala, Sweden), zVAD-fmk (caspase-3 inhibitor) from the Peptide In-stitute (Osaka, Japan), 2-methyl-6-methoxyphenylethynylimidazo-pyrazynone (MPEC) from ATTOCorp. (Osaka, Japan), 5,5 -dithiobis-2-nitrobenzoic acid (DTNB) from DOJINDO (Kumamoto, Japan),N -(Biotinoyl)-N 9-(iodoacetyl)ethylenediamine (BIAM) from Invitrogen (Carlsbad, CA, USA), mousemonoclonal anti-human band 3 antibody and horseradish peroxidase (HRP)-conjugated streptavidinfrom Sigma (St. Louis, MO, USA), rabbit polyclonal anti-human band 3 antibody from Santa CruzBiotechnology (Santa Cruz, CA, USA), and the Protein Assay kit from Bio-Rad (Hercules, CA, USA).
Ginsenosides Rb1, Rb2, Rc, Rd, Re, Rg1, Rg2, Rg3, Rh1, Rh2 and Ro were isolated and purified
from the crude saponin fraction of the rhizome of Panax Ginseng C.A. Meyer, Korea Red Ginseng, by
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
repeated-column chromatography on silica gel with CHCl3–MeOH–H2O (65:35:10) and on octadecylsi-lyl (ODS) silica with MeOH–H2O (1:1–7:3) [24,31]. The purity of each of the 11 ginsenosides used inthis study was more than 99.99%, as determined by high-performance liquid chromatography.
Heparinized venous blood was obtained, after written informed consent, from healthy donors, and
immediately centrifuged. After a careful removal of plasma and buffy coat, erythrocytes were purifiedby three cycles of resuspension and washing with isotonic HEPES-buffered saline (HBS) containing100 mg/dl glucose at 4◦C, and resuspended at 40% hematocrit in HBS for further analysis. 2.4. Exposure of erythrocytes to oxidative stress
The erythrocyte suspensions of 10% hematocrit were treated with 0.5 mM FeSO4 with 5 mM ascorbate
(final concentrations) at 37◦C for 1 h with gentle shaking [7]. The incubation was carried out in thepresence or absence of 0.05 mg/ml total ginsenosides or 0.01 mg/ml of one of the ginsenoside fractions(final concentrations). The contents of Rg2 and Rh1 in 0.05 mg/ml total ginsenosides are calculated as0.0001 and 0.0007 mg/ml, respectively [24].
The viscosity of erythrocyte suspensions of 45% hematocrit was measured at 25◦C with a cone–plate
viscometer at shear rates from 19 to 376 s−1. The deformability of erythrocytes was measured with ahigh-shear rheoscope under 2.36 Pa shear stress. The measurement was performed at 0.3% hematocritin isotonic HBS containing 25% Dextran T-40, at 25◦C. The degree of ellipsoidal deformation of ery-throcytes was expressed as the deformation index DI = (L − S)/(L + S), where L and S were the majorand minor radii of the deformed cells, respectively [27].
Erythrocyte suspension (250 µl at 40% hematocrit) was lysed on ice with 1.75 ml of hypotonic PBS
(20 mOsm), pH 7.4, and centrifuged for 10 min at 4◦C at 10,000 × g. After removal of clear supernatant,the sediment was resuspended in ice-cold hypotonic PBS and centrifuged again. The washing procedurewas repeated three times to obtain the membrane ghosts. 2.7. Thiobarbituric acid reactive substances (TBARS) assay
Lipid peroxidation products formed from the oxidative breakdown of polyunsaturated fatty acids were
measured in erythrocyte ghost membranes using the TBARS assay as described previously [7]. Briefly,0.15 ml of the ghosts sample solubilized with 1 g/dl SDS was combined with 0.45 ml of a solutioncontaining 15 g/dl TCA, 0.375 g/dl TBA, 0.25 M HCl and 0.015 g/dl BHT. The solution was heated for15 min in a boiling water bath. After cooling, the flocculent precipitates were removed by centrifugationat 1000 × g for 5 min and the light absorbance of the sample was measured at 535 nm. The TBARSconcentration was calculated using the extinction coefficient 1.56 × 105 M−1 and expressed in nM ofTBARS per gram hemoglobin. K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes2.8. Measurement of antioxidant activity
MPEC stock solution (0.075 M) in ethanol was stored at −40◦C. To quantify the antioxidant activity
of each ginsenosides, the assay was carried out by using the MPEC according to the manufacturer’sinstructions. Briefly, 0.0025% hydrogen peroxide was added to the solution containing 10 mM MPEC(final concentration) and one of the following substances (final concentrations): 0.5 mg/ml vitamin C,0.2 mg/ml vitamin E, 0.15 mg/ml total ginsenosides, or 0.03 mg/ml of one of the following ginsenosides:Rb1, Rg2 or Rh1. Subsequently, the light emission was measured with luminometer (Model TD-20/20,Turner Designs). 2.9. Measuring the concentration of SH-groups on the erythrocyte membrane
Erythrocyte ghosts (0.1 ml) solubilized with 1 g/dl SDS were mixed with 0.9 ml of 100 mM sodium
phosphate (pH 7.4) containing 0.1 mM EDTA. Then, 5,5 -dithiobis-2-nitrobenzoic acid (DTNB) wasadded at a final concentration of 1 mM. After incubation at 37◦C for 15 min, the absorbance at 412 nmwas measured. The concentration of SH-groups was calculated using molar extinction coefficient of15,200. Total protein concentration was determined with bovine serum albumin as a standard by themethod of Bradford et al. [3] using the Protein Assay kit (Bio-Rad) according to the manufacturer’sprotocol. The ratio of SH-group to total protein relative to control was used as an index of antioxidantactivity. 2.10. SDS-polyacrylamide gel electrophoresis (PAGE)
The erythrocyte membrane ghosts were subjected to SDS-PAGE using 7.5% polyacrylamide gel and
stained with 0.025% Coomassie brilliant blue. 2.11. Labeling of SH-group with BIAM
Erythrocyte ghosts were suspended in hypotonic PBS (20 mOsm), pH 7.4. SH-groups in the erythro-
cyte ghosts were labeled with 100 µM BIAM for 15 min at room temperature in the dark. The suspensionwas centrifuged for 10 min at 4◦C at 10,000×g. After removal of clear supernatant, the sediment was re-suspended in ice-cold hypotonic PBS and centrifuged again. The washing procedure was repeated threetimes. The final sediment was lysed with lysing buffer containing 20 mM 2-mercaptoethanol, 50 mMTris–HCl, pH 8.0, 20 mM EDTA, 1% SDS, and 100 mM NaCl, followed by boiling for 5 min and SDS-PAGE. The separated proteins were transferred to a PVDF membrane. Proteins labeled with BIAM weredetected with HRP-conjugated streptavidin using an ECL Plus Western Blotting Detection System (GEHealthcare Bio-Sciences).
Group differences were analyzed with one-way analysis of variance using SPSS 6.1J software. The
critical significance level was set at p < 0.05. Post-test analyses using a paired t-test were carried out. All data are presented as mean ± SD. K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes3. Results 3.1. Screening for the components of Panax ginseng which attenuate the oxidation-induced impairmentof erythrocyte membrane properties
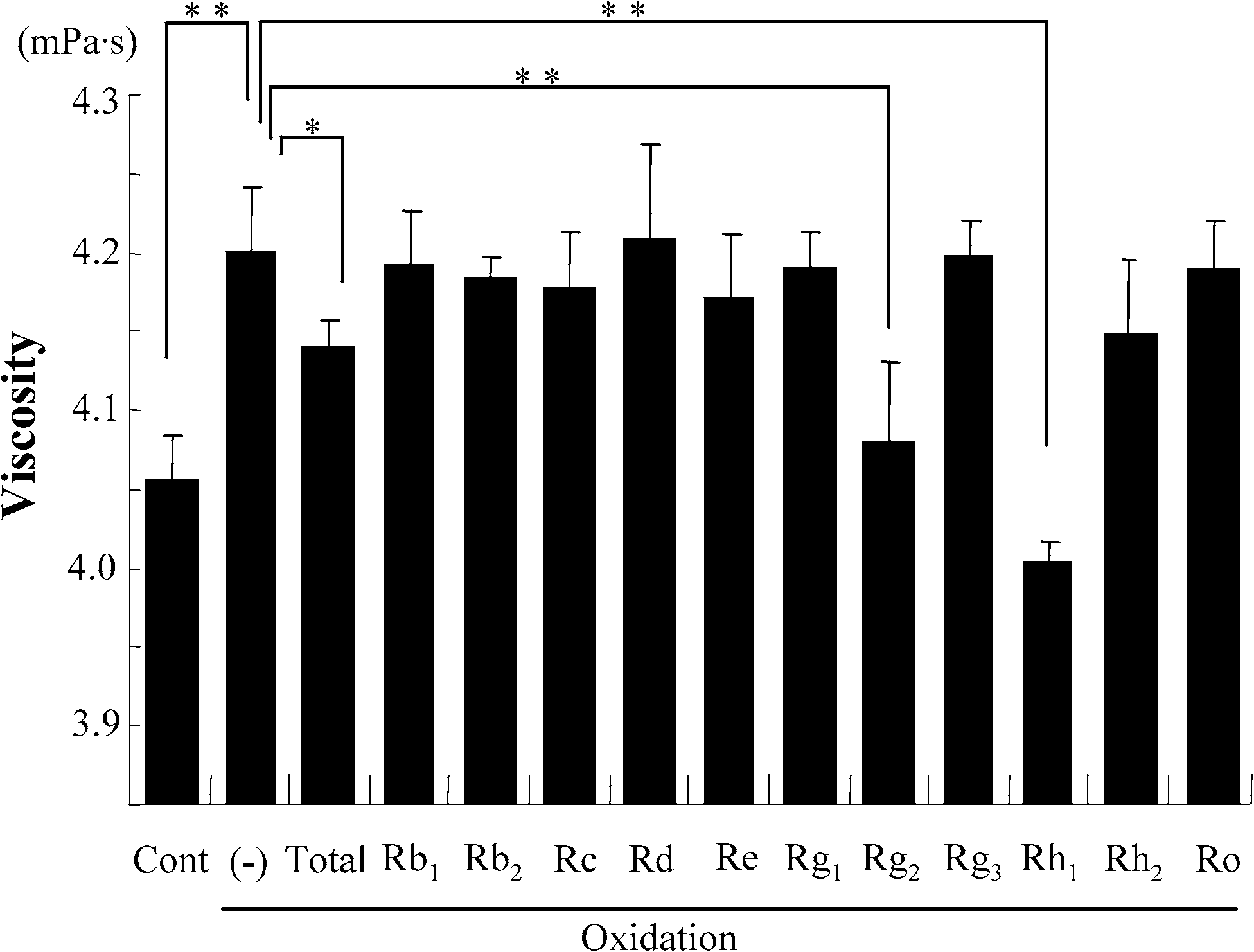
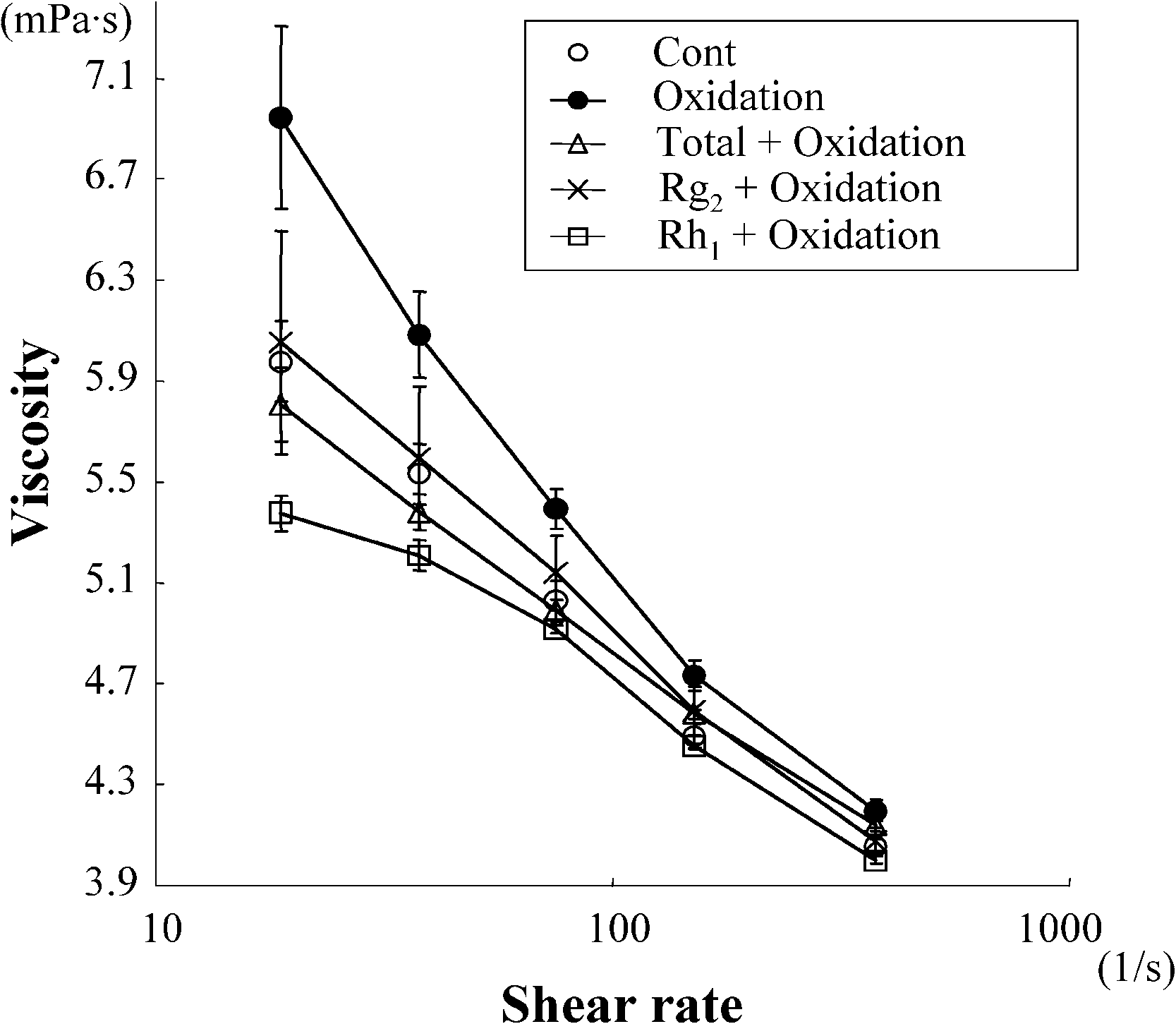
Exposure of the erythrocyte suspension to oxidative stress resulted in a significant increase of viscosity
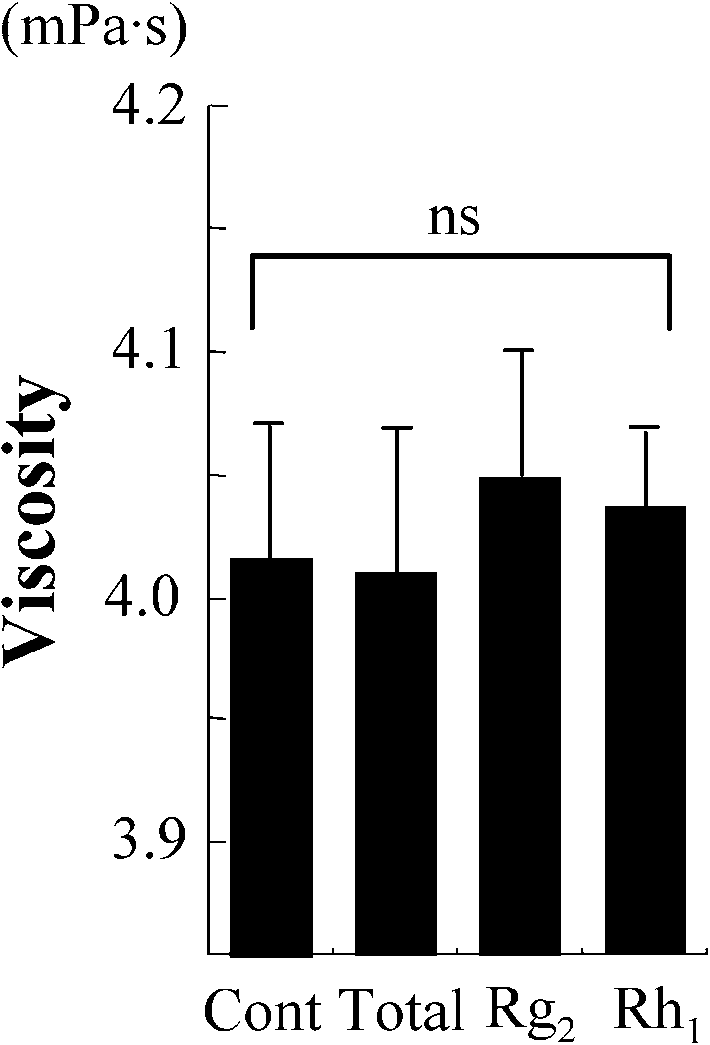
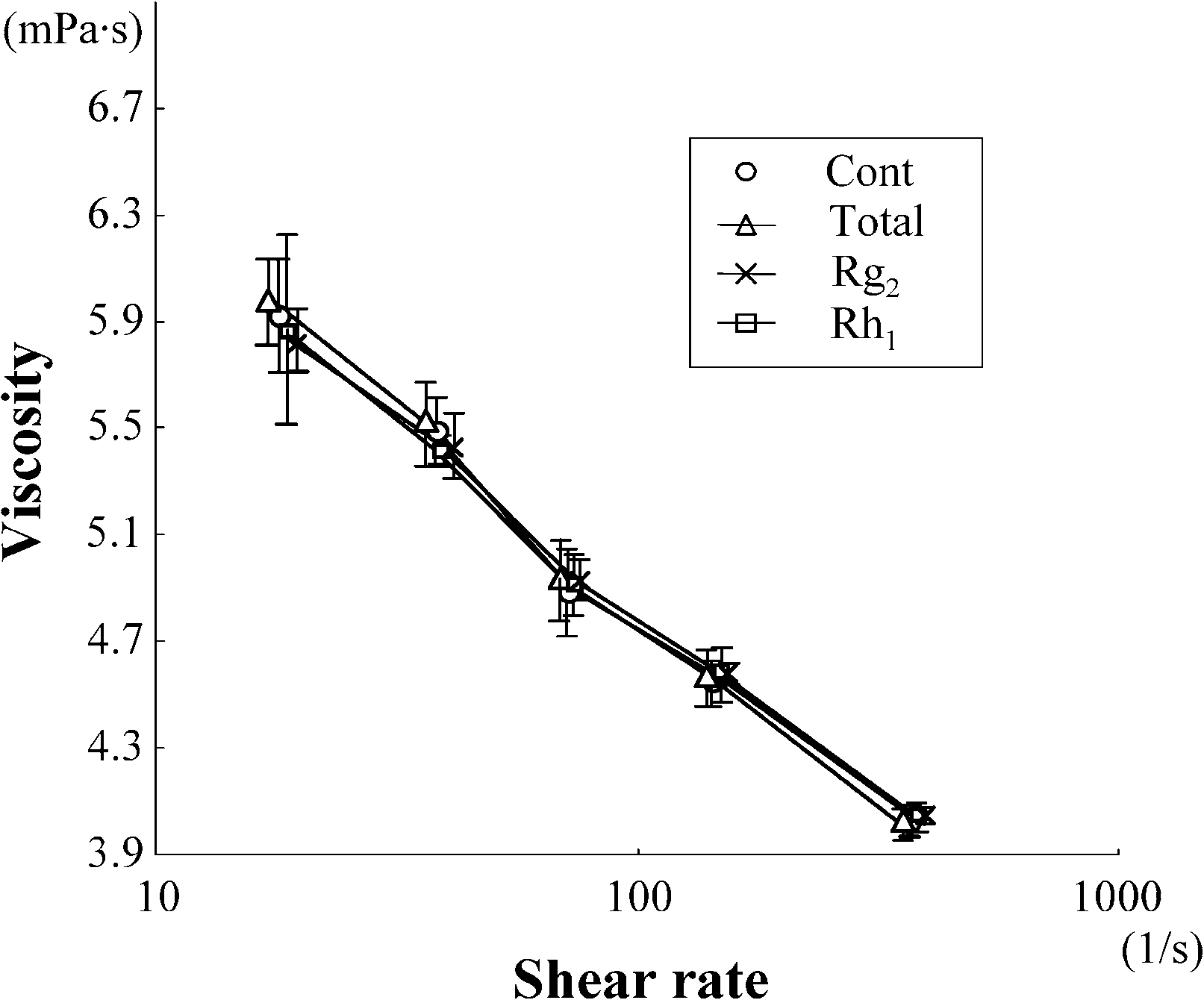
measured at high shear rate (380 s−1). Treatment with total ginsenosides attenuated this increase ofviscosity. We first screened each of ginsenosides using this attenuating effect as an index of beneficialactivity, and identified Rg2 and Rh1 as the components capable of preventing the oxidative stress-inducedincrease in viscosity (Fig. 1(A)). As shown in Fig. 1(B), both Rg2 and Rh1 attenuated the oxidation-induced increase of viscosity over all of the used range of shear rate. Treatment of the erythrocytesuspension with total ginsenosides, Rg2 and Rh1 without exposing to oxidative stress did not alter theviscosity (Fig. 1(C and D)). These results suggest that these active agents attenuate the oxidative stress-induced increase of viscosity without changing the intrinsic properties of the erythrocytes.
We next tested the effect of these components on erythrocyte deformability. The exposure of the
erythrocyte suspension to oxidative stress resulted in a significant decrease of deformability. Treatmentwith Rg2 or Rh1 from Panax ginseng, as well as with total ginsenosides, significantly inhibited thedecrease (Fig. 2). No hemolysis was observed in any of the experiments. 3.2. Rg2 and Rh1 from Panax ginseng do not show antioxidant activity
The antioxidant activity of the identified active components of Panax ginseng was measured using
MPEC. Vitamin C, vitamin E, total ginsenosides and ginsenoside Rb1, all significantly inhibited thelight emission induced by hydrogen peroxide. However, Rg2 and Rh1, previously identified as activecomponents, did not inhibit the light emission significantly, even when tested at much higher concentra-tions than the total ginsenosides (Fig. 3). 3.3. Ginsenosides from Panax ginseng do not protect erythrocytes from peroxidation of membrane
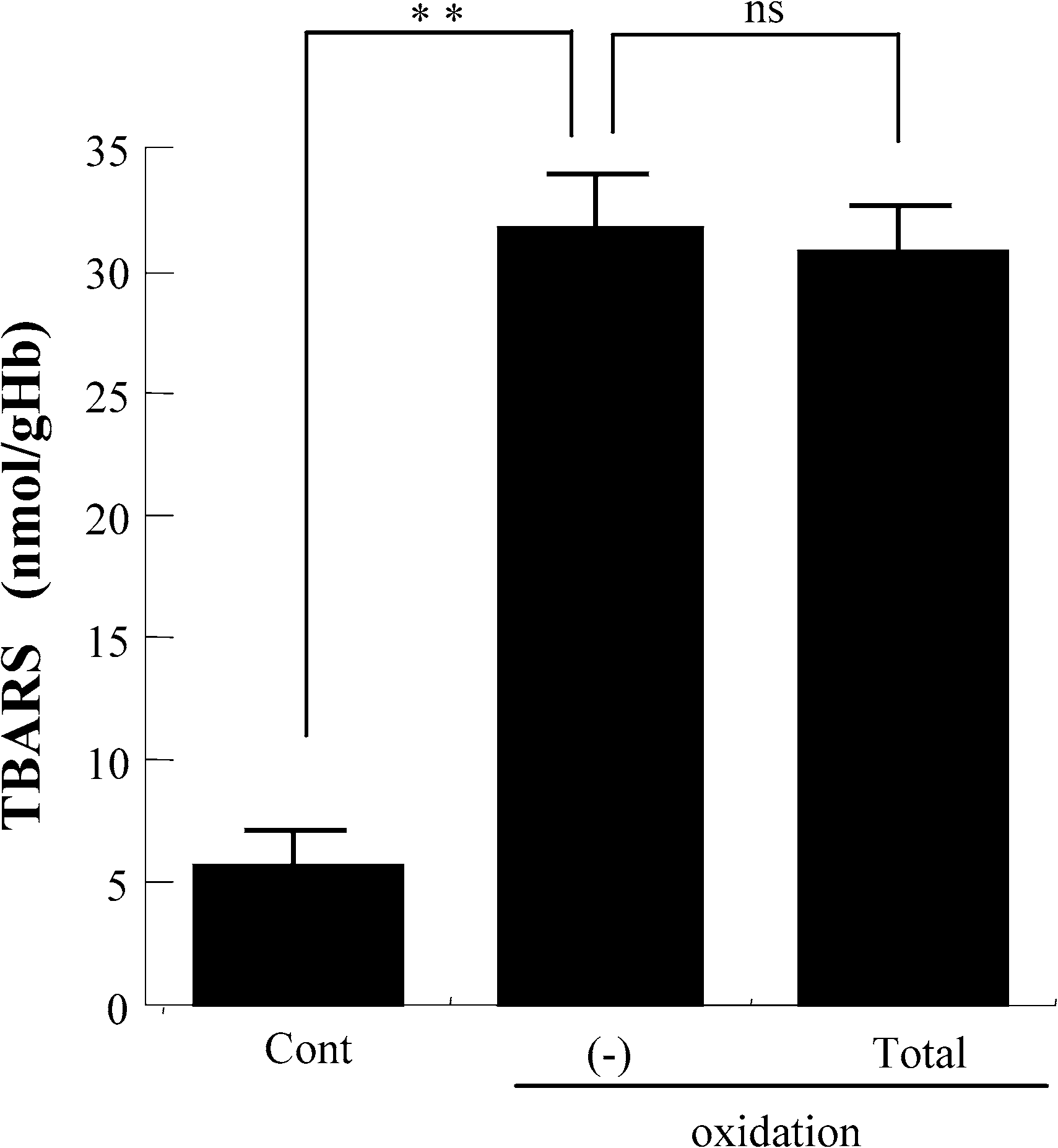
As we previously reported, exposure of erythrocytes suspension to oxidative stress resulted in a dose-
dependent oxidation of membrane lipids reflected by the formation of TBARS [27]. In the present study,even treatment with total ginsenosides did not affect TBARS level in erythrocytes exposed to oxidativestress (Fig. 4). This finding suggests that ginsenosides protect erythrocyte membrane properties fromoxidative stress via a mechanism other than inhibition of oxidation of membrane lipids. 3.4. Rg2 and Rh1 from Panax ginseng prevent the decrease in SH-groups of band 3 protein
The exposure of the erythrocyte suspension to oxidative stress resulted in a significant decrease of
SH-groups in the membrane. Treatment with either total ginsenosides, or with the active components,Rg2 or Rh1, significantly attenuated the decrease. In contrast, Rb1 showed no beneficial effect on theamount of SH-groups in the erythrocyte membranes (Fig. 5(A)).
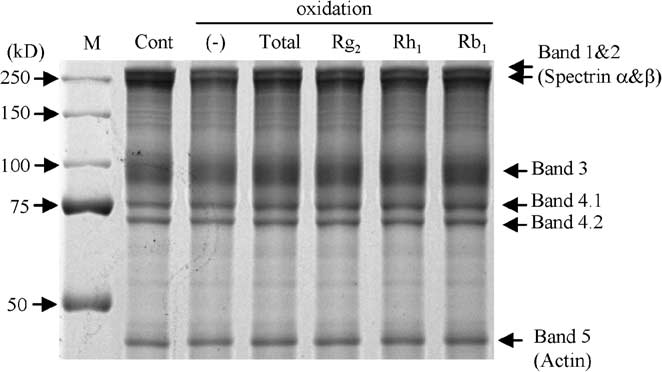
Although the total amounts of protein loaded to each lane were same, the SDS-PAGE gel stained with
Coomassie brilliant blue (Fig. 5(B)), showed that the amount of bands 1 and 2 (spectrins), and band 3appeared to decrease a little under the oxidative conditions we used. As we previously reported [27], thisphenomenon is probably due to oxidation-induced activation of caspase-3, a proteolytic enzyme which
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 1. Screening for the components of Panax ginseng which attenuate the oxidation-induced increase in viscosity of erythro-cyte suspensions. Data represent mean ± SD (n = 10). Total: total ginsenosides; ∗p < 0.05; ∗∗p < 0.01. (A) The viscosityof erythrocyte suspensions unexposed (Cont) or exposed to oxidative stress (Oxidation) in the presence or absence of eachginsenoside, measured under high shear rate. (B) The viscosity of erythrocyte suspensions unexposed (Cont) or exposed to ox-idative stress (Oxidation) in the presence or absence of Rg2 or Rh1, measured over all of the range of shear rates used. (C) Theviscosity of erythrocyte suspensions unexposed to oxidative stress in the presence or absence (Cont) of total ginsenosides, Rg2or Rh1, measured under high shear rate. ns: no significant difference. (D) The viscosity of erythrocyte suspensions unexposedto oxidative stress in the presence or absence (Cont) of total ginsenosides, Rg2 or Rh1, measured over all of the range of shearrate used.
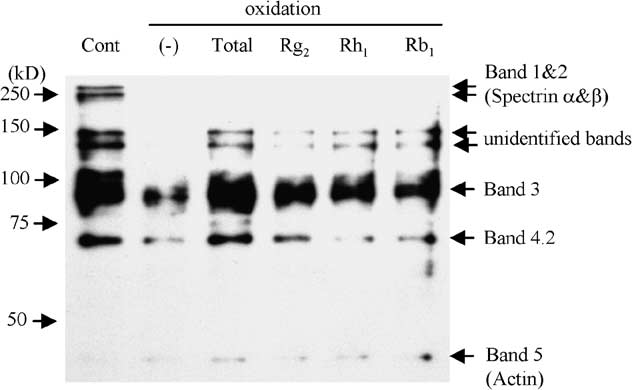
cleaves the spectrin and band 3 in the erythrocyte membrane [20,30]. There were several SH-rich bandsin the control unexposed to oxidative stress (Fig. 5(C), lane 1), which were assumed to be bands 1, 2, 3and 4.2. Moreover, there were two unidentified SH-rich bands around 140 and 130 kDa. When erythro-cytes were exposed to oxidative stress, the content of the SH-groups in each band dramatically decreased(lane 2). Interestingly, treatment with Rg2 or Rh1 from Panax ginseng, as well as with total ginsenosides,significantly attenuated the decrease in SH-groups of band 3, but not of bands 1 and 2 (lanes 3–5). Totalginsenosides and Rg2 appeared to prevent the decrease in SH-groups of band 4.2, whereas Rh1 did not. Rb1 did not appear to attenuate the decrease at all (lane 6). Subsequent immunoprecipitation with mon-
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 2. Protective effect of Rg2 and Rh1 on oxidation-induced impairment of erythrocyte membrane deformability. Cont: controlunexposed to oxidative stress; total: total ginsenosides; ∗p < 0.05; ∗∗p < 0.01. Values are mean ± SD (n = 10) of thedeformation index.
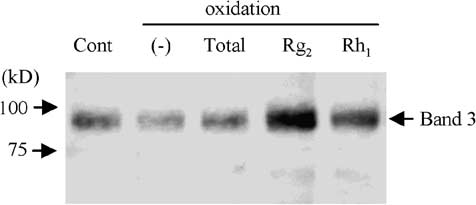
oclonal anti-band 3 antibody, followed by SDS-PAGE and staining with HRP-conjugated streptavidinconfirmed that Rg2 and Rh1 attenuated the decrease in SH-groups of band 3 protein (Fig. 5(D)). 4. Discussion
This study was carried out in order to investigate the effects of ginsenosides on oxidative stress-
induced impairment of the rheological properties of erythrocytes. In addition, we attempted to identifythe active components of Panax ginseng extract and to clarify the mechanisms by which they protecterythrocytes from oxidative injury. Low-level oxidative stress used here resulted in an increased erythro-cyte suspension viscosity and a reduced deformability of the exposed erythrocytes. Treatment with totalginsenosides significantly attenuated the oxidation-induced elevation of the viscosity of the erythrocytesuspensions measured from low to high shear rates. It also significantly attenuated the impairment of theerythrocyte membrane deformability. Subsequent analyses identified ginsenosides Rg2 and Rh1 as theactive components of the Panax ginseng extract. These two ginsenosides effectively inhibited the ox-idative stress-induced impairment of the rheological properties of erythrocytes whereas other examinedginsenosides had no effect.
Rg2 and Rh1 are minor components of saponin, belonging to protopanaxatriol group. The contents of
total ginsenosides, Rg2 and Rh1 are 24.057, 0.068 and 0.340 mg, respectively, in 1 g Panax ginseng [24]. K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 3. Measurement of antioxidant activity. Data expressed as the ratio of emission units relative to control. The values repre-sented by bars are mean ± SD (n = 20). Cont: control not exposed to oxidative stress; VC: vitamin C; VE: vitamin E; total:total ginsenosides; ∗∗p < 0.01; ns: no significant difference.
Fig. 4. The effect of total ginsenosides on oxidation of erythrocyte membrane lipids. Cont: control not exposed to oxidativestress; total: total ginsenosides; ∗∗p < 0.01; ns: no significant difference. Values are mean ± SD (n = 10) of TBARS,membrane lipid oxidation expressed as nM of TBARS per gram hemoglobin. K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 5. SH-group content in erythrocyte membrane. Cont: control not exposed to oxidative stress; total: total ginsenosides. (A) Measurement of SH-group content in erythrocyte membrane exposed to oxidative stress in the presence or absence ofginsenosides. Data represent the ratio of (SH-group/total protein) relative to control. The values represented by the bars aremean ± SD (n = 10); p < 0.01. (B) SDS-PAGE gel stained with Coomassie brilliant blue. (C) Erythrocyte membrane proteinslabeled with BIAM were subjected to SDS-PAGE. SH-groups were detected with HRP-conjugated streptavidin and ECL PlusWestern Blotting Detection System. (D) Erythrocyte membrane proteins labeled with BIAM were immunoprecipitated withanti-band 3 antibody, subjected to SDS-PAGE, followed by staining with HRP-conjugated streptavidin and ECL Plus WesternBlotting Detection System.
Both Rg2 and Rh1 have hydroxyl group at positions-3 and -20, the only structural difference beingposition-6, where Rg2 has an α-L-rhamnopyranosyl-(1→2)-β-D-glucopyranosyl group, while Rh1 has aβ-D-glucopyranosyl group. Very little was reported so far about the effects and the mechanisms of thesetwo minor components. Among the existing reports, recent studies demonstrated the anti-apoptotic effectof ginsenoside Rg2, which protects NIH3T3 cells against ultraviolet B-induced apoptosis by increasingDNA repair [10] and prevents memory impairment via anti-apoptosis in a rat model with vascular de-mentia [33]. Moreover, Rg2 has been shown to protect neuronal cells against glutamate-induced neuronalinjury by inhibiting excessive Ca2+ influx, reducing the lipid peroxidation and down-regulating the ex-pression of the pro-apoptotic factor calpain II and caspase-3 [17]. As for ginsenoside Rh1, it was reportedto possess anti-allergic and anti-inflammatory activities [21].
In this study, the mechanisms by which Rg2 and Rh1 protect the rheological functions of erythrocytes
do not appear to be associated with the inhibition of membrane lipid peroxidation, because even the
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
total ginsenosides did not attenuate the formation of lipid peroxidation products from the oxidativebreakdown of polyunsaturated fatty acids in the erythrocyte membrane. The existing data suggest thatthe mechanism of the protective effect of ginsenosides against oxidative stress may be related to theirantioxidant activity [12,13,15,19,32]. Thus, we tested the antioxidant activity of Rg2 and Rh1 using theMPEC reaction kit. Contrary to our expectations, the antioxidant activities of Rg2 and Rh1 were notsignificantly higher than that of the control, while those of the total ginsenosides and Rb1, as well asvitamin C and vitamin E, were significantly higher. Theoretically, the MPEC kit, where 2-methyl-6-methoxyphenylethynylimidazopyrazynone (MPEC) reacts with superoxide anion in the aqueous phaseand emits luminescence, measures the antioxidant activity only in the aqueous phase. Because Rg2 andRh1 are both lipid-soluble, water-insoluble molecules, they may display their antioxidant activity inthe lipid phase rather than in the aqueous phase. They can intercalate into the erythrocyte membrane,and may take effect there. Therefore, our results of the MPEC assay do not necessarily contradict thepreviously implicated antioxidant activity of ginsenosides.
Regarding the mechanism of action, Rg2 and Rh1 appeared to inhibit the oxidation of the SH-group
in the cysteine residue of the erythrocyte membrane protein. The oxidation-induced decrease of theSH-group in the erythrocyte membrane appeared to be related mainly to bands 1 and 2 (spectrins αand β), band 3 (anion exchanger-1) and band 4.2. Comparing different membrane proteins, Rg2 andRh1 attenuated the decrease of the SH-group particularly in band 3 protein. The erythrocyte membranecytoskeletal protein band 3 is a structural protein that confers stability and flexibility to erythrocytes. Itplays an important role in maintaining the erythrocytes’ biconcave shape and their resilience to shearforce [9]. It is a 95-kDa glycoprotein, accounting for approximately 25% w/w of the total protein inthe erythrocyte membrane. In this protein, there are five SH-containing cysteine residues, two of whichare in the N-terminal domain, two in the transmembrane domain, and one is in C-terminal domain [1]. Its N-terminal cytoplasmic domain binds to cytoskeletal protein spectrin via ankyrin [4]. The 201st and317th cysteine residues in the N-terminal domain of band 3 are very important in the formation ofband 3 dimer and binding to ankyrin [28]. Furthermore, its C-terminal transmembrane domain medi-ates the HCO−/Cl− anion exchange [29], leading to the O
2/CO2 exchange in erythrocytes. Oxidation of
SH-groups in this domain leads to impaired phosphate transport, which results in decreased deformabil-ity of erythrocytes [22].
In the present study, a relatively weak oxidative stress was used, which did not lead to strong degra-
dation of band 3, but was capable of impairing erythrocyte membrane functions, probably via depletionof SH-groups. Under these conditions, the number of SH-groups in band 3 appeared to decrease muchmore than band 3 protein itself. These results suggest that with increasing scale of oxidative damage,SH-group depletion seems to precede the degradation of band 3, which occurs at higher levels of ox-idative stress and requires activation of caspase-3 [27]. The mechanism by which oxidation decreasesSH-groups in the erythrocyte membrane was shown to be independent of caspase-3 activity, since treat-ment with caspase inhibitor (zVAD-fmk) did not attenuate the oxidation-induced decrease of SH-groups(data not shown). According to the results obtained, the protective effect of Rg2 and Rh1 was not relatedto the inhibition of caspase-3 activation. Thus, although these active compounds of Panax ginseng ex-tract do not prevent band 3 degradation, they effectively inhibit the oxidation of the SH-groups of band 3protein.
Taken together, the present work has shown that the oxidative damage of band 3 protein can disturb
membrane–cytoskeletal interactions and anion exchange, leading to an increased erythrocyte suspensionviscosity and a reduced deformability of the exposed erythrocytes. Treatment of erythrocyte suspensions
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
with Rg2 and Rh1 protects the rheological functions of the erythrocyte by inhibiting the oxidation ofSH-groups in band 3 protein. Acknowledgement
This work was supported by grant-in-aid for scientific research from the Ministry of Education, Cul-
ture, Sports, Science and Technology of Japan, and a grant from Hongsam.com, Ltd. (Seoul, Korea). References
[1] S.L. Alper, Molecular physiology of SLC4 anion exchangers, Exp. Physiol. 91 (2006), 153–161. [2] A.S. Attele, J.A. Wu and C.S. Yuan, Ginseng pharmacology: multiple constituents and multiple actions, Biochem. Phar- macol. 58 (1999), 1685–1693.
[3] M.M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle
of protein-dye binding, Anal. Biochem. 72 (1976), 248–254.
[4] J.R. Casey and R.A. Reithmeier, Anion exchangers in the red cell and beyond, Biochem. Cell Biol. 76 (1998), 709–713. [5] S.W. Chen, X.H. Li, K.H. Ye, Z.F. Jiang and X.D. Ren, Total saponins of Panax notoginseng protected rabbit iliac artery
against balloon endothelial denudation injury, Acta Pharmacol. Sin. 25 (2004), 1151–1156.
[6] T.W. Chung and C.P. Ho, Changes in viscosity of low shear rates and viscoelastic properties of oxidative erythrocyte
suspensions, Clin. Hemorheol. Microcirc. 21 (1999), 99–103.
[7] I. Cicha, N. Tateishi, Y. Suzuki and N. Maeda, Rheological changes in human red blood cells under oxidative stress,
Pathophysiology 6 (1999), 103–110.
[8] F. He, R. Guo, S.L. Wu, M. Sun and M. Li, Protective effects of ginsenoside Rb1 on human umbilical vein endothelial
cells in vitro, J. Cardiovasc. Pharmacol. 50 (2007), 314–320.
[9] D.G. Jay, Role of band 3 in homeostasis and cell shape, Cell 86 (1996), 853–854.
[10] S.J. Jeong, S.H. Han, D.Y. Kim, J.C. Lee, H.S. Kim, B.H. Kim, J.S. Lee, E.H. Hwang and J.K. Park, Effects of mRg2,
a mixture of ginsenosides containing 60% Rg2, on the ultraviolet B-induced DNA repair synthesis and apoptosis in NIH3T3 cells, Int. J. Toxicol. 26 (2007), 151–158.
[11] C.S. Kim, J.B. Park, K.J. Kim, S.J. Chang, S.W. Ryoo and B.H. Jeon, Effect of Korea red ginseng on cerebral blood flow
and superoxide production, Acta Pharmacol. Sin. 23 (2002), 1152–1156.
[12] K.T. Kim, K.M. Yoo, J.W. Lee, S.H. Eom, I.K. Hwang and C.Y. Lee, Protective effect of steamed American ginseng
(Panax quinquefolius L.) on V79-4 cells induced by oxidative stress, J. Ethnopharmacol. 111 (2007), 443–450.
[13] Y.K. Kim, Q. Guo and L. Packer, Free radical scavenging activity of red ginseng aqueous extracts, Toxicology 172 (2002),
[14] Y. Kimura, M. Sumiyoshi, K. Samukawa, N. Satake and M. Sakanaka, Facilitating action of asiaticoside at low doses on
burn wound repair and its mechanism, Eur. J. Pharmacol. 584 (2008), 415–423.
[15] D.D. Kitts, A.N. Wijewickreme and C. Hu, Antioxidant properties of a North American ginseng extract, Mol. Cell.Biochem. 203 (2000), 1–10.
[16] H.U. Lee, E.A. Bae, M.J Han, N.J. Kim and D.H. Kim, Hepatoprotective effect of ginsenoside Rb1 and compound K on
tert-butyl hydroperoxide-induced liver injury, Liver Int. 25 (2005), 1069–1073.
[17] N. Li, B. Liu, D.E. Dluzen and Y. Jin, Protective effects of ginsenoside Rg2 against glutamate-induced neurotoxicity in
PC12 cells, J. Ethnopharmacol. 111 (2007), 458–463.
[18] X. Liu, W. Qin and D. Yin, Biochemical relevance between oxidative/carbonyl stress and elevated viscosity of erythrocyte
suspensions, Clin. Hemorheol. Microcirc. 31 (2004), 149–156.
[19] Z.Q. Liu, X.Y. Luo, G.Z. Liu, Y.P. Chen, Z.C. Wang and Y.X. Sun, In vitro study of the relationship between the structure
of ginsenoside and its antioxidative or prooxidative activity in free radical induced hemolysis of human erythrocytes, J. Agric. Food Chem. 51 (2003), 2555–2558.
[20] D. Mandal, V. Baudin-Creuza, A. Bhattacharyya, S. Pathak, J. Delaunay, M. Kundu and J. Basu, Caspase 3-mediated
proteolysis of the N-terminal cytoplasmic domain of the human erythroid anion exchanger 1 (band 3), J. Biol. Chem. 278 (2003), 52551–52558.
[21] E.K. Park, M.K. Choo, M.J. Han and D.H. Kim, Ginsenoside Rh1 possesses antiallergic and anti-inflammatory activities,
Int. Arch. Allergy Immunol. 133 (2004), 113–120.
[22] S.S. Roy, G. Sen and T. Biswas, Role of sulfhydryl groups in band 3 in the inhibition of phosphate transport across
erythrocyte membrane in visceral leishmaniasis, Arch. Biochem. Biophys. 436 (2005), 121–127. K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
[23] M. Sakanaka, P. Zhu, B. Zhang, T.C. Wen, F. Cao, Y.J. Ma, K. Samukawa, N. Mitsuda, J. Tanaka, M. Kuramoto, H. Uno
and R. Hata, Intravenous infusion of dihydroginsenoside Rb1 prevents compressive spinal cord injury and ischemic brain damage through upregulation of VEGF and Bcl-XL, J. Neurotrauma 24 (2007), 1037–1054.
[24] K. Samukawa, H. Yamashita, H. Matsuda and M. Kubo, Simultaneous analysis of ginsenosides of various ginseng radix
by HPLC, Yakugaku Zasshi 115 (1995), 241–249.
[25] K. Sun, C.S. Wang, J. Guo, Y. Horie, S.P. Fang, F. Wang, Y.Y. Liu, L.Y. Liu, J.Y. Yang, J.Y. Fan and J.Y. Han, Protective
effects of ginsenoside Rb1, ginsenoside Rg1, and notoginsenoside R1 on lipopolysaccharide-induced microcirculatory disturbance in rat mesentery, Life Sci. 81 (2007), 509–518.
[26] J. Sung, K.H. Han, J.H. Zo, H.J. Park, C.H. Kim and B.H. Oh, Effects of red ginseng upon vascular endothelial function
in patients with essential hypertension, Am. J. Clin. Med. 28 (2000), 205–216.
[27] Y. Suzuki, N. Ohkubo, M. Aoto, N. Maeda, I. Cicha, T. Miki and N. Mitsuda, Participation of caspase-3-like protease in
oxidation-induced impairment of erythrocyte membrane properties, Biorheology 44 (2007), 179–190.
[28] B.J. Thevenin, B.M. Willardson and P.S. Low, The redox state of cysteines 201 and 317 of the erythrocyte anion exchanger
is critical for ankyrin binding, J. Biol. Chem. 264 (1989), 15886–15892.
[29] D.N. Wang, Band 3 protein: structure, flexibility and function, FEBS Lett. 346 (1994), 26–31. [30] K.K. Wang, R. Posmantur, R. Nath, K. McGinnis, M. Whitton, R.V. Talanian, S.B. Glantz and J.S. Morrow, Simultaneous
degradation of alphaII- and betaII-spectrin by caspase 3 (CPP32) in apoptotic cells, J. Biol. Chem. 273 (1998), 22490– 22497.
[31] B. Zhang, R. Hata, P. Zhu, K. Sato, T.C. Wen, L. Yang, H. Fujita, N. Mitsuda, J. Tanaka, K. Samukawa, N. Maeda and
M. Sakanaka, Prevention of ischemic neuronal death by intravenous infusion of a ginseng saponin, ginsenoside Rb(1), that upregulates Bcl-x(L) expression, J. Cerebr. Blood Flow Metab. 26 (2006), 708–721.
[32] D. Zhang, T. Yasuda, Y. Yu, P. Zheng, T. Kawabata, Y. Ma and S. Okada, Ginseng extract scavenges hydroxyl radical and
protects unsaturated fatty acids from decomposition caused by iron-mediated lipid peroxidation, Free Radic. Biol. Med. 20 (1996), 145–150.
[33] G. Zhang, A. Liu, Y. Zhou, X. San, T. Jin and Y. Jin, Panax ginseng ginsenoside-Rg2 protects memory impairment via
anti-apoptosis in a rat model with vascular dementia, J. Ethnopharmacol. 115 (2008), 441–444.
nineteen years on the job Part 2 of a 2-part article by Lindy Dunagan-Groger [Volume 12 Number 2 (June 2002) features part 1 of this article] n the year that Michael turned 18 I transitioned into a new set of I am an organized documentation- issues — those of parenting and being person; I have kept all those papers and a young adult. This included officially obtaining disabili
REMPLACE L’APPENDICE A DU CAMO 1999 CODE ANTIDOPAGE DU MOUVEMENT OLYMPIQUE APPENDICE A CLASSES DE SUBSTANCES INTERDITES ET MÉTHODES INTERDITES 1 avril 2000 CLASSES DE SUBSTANCES INTERDITES Les substances interdites appartenant à la classe (A) comprennent les exemples suivants : amineptine, amiphénazole, amphétamines, bromantan, caféine*, carphédon, cocaïne, éphédrines
 K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 1. Screening for the components of Panax ginseng which attenuate the oxidation-induced increase in viscosity of erythro-cyte suspensions. Data represent mean ± SD (n = 10). Total: total ginsenosides; ∗p < 0.05; ∗∗p < 0.01. (A) The viscosityof erythrocyte suspensions unexposed (Cont) or exposed to oxidative stress (Oxidation) in the presence or absence of eachginsenoside, measured under high shear rate. (B) The viscosity of erythrocyte suspensions unexposed (Cont) or exposed to ox-idative stress (Oxidation) in the presence or absence of Rg2 or Rh1, measured over all of the range of shear rates used. (C) Theviscosity of erythrocyte suspensions unexposed to oxidative stress in the presence or absence (Cont) of total ginsenosides, Rg2or Rh1, measured under high shear rate. ns: no significant difference. (D) The viscosity of erythrocyte suspensions unexposedto oxidative stress in the presence or absence (Cont) of total ginsenosides, Rg2 or Rh1, measured over all of the range of shearrate used.
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 1. Screening for the components of Panax ginseng which attenuate the oxidation-induced increase in viscosity of erythro-cyte suspensions. Data represent mean ± SD (n = 10). Total: total ginsenosides; ∗p < 0.05; ∗∗p < 0.01. (A) The viscosityof erythrocyte suspensions unexposed (Cont) or exposed to oxidative stress (Oxidation) in the presence or absence of eachginsenoside, measured under high shear rate. (B) The viscosity of erythrocyte suspensions unexposed (Cont) or exposed to ox-idative stress (Oxidation) in the presence or absence of Rg2 or Rh1, measured over all of the range of shear rates used. (C) Theviscosity of erythrocyte suspensions unexposed to oxidative stress in the presence or absence (Cont) of total ginsenosides, Rg2or Rh1, measured under high shear rate. ns: no significant difference. (D) The viscosity of erythrocyte suspensions unexposedto oxidative stress in the presence or absence (Cont) of total ginsenosides, Rg2 or Rh1, measured over all of the range of shearrate used.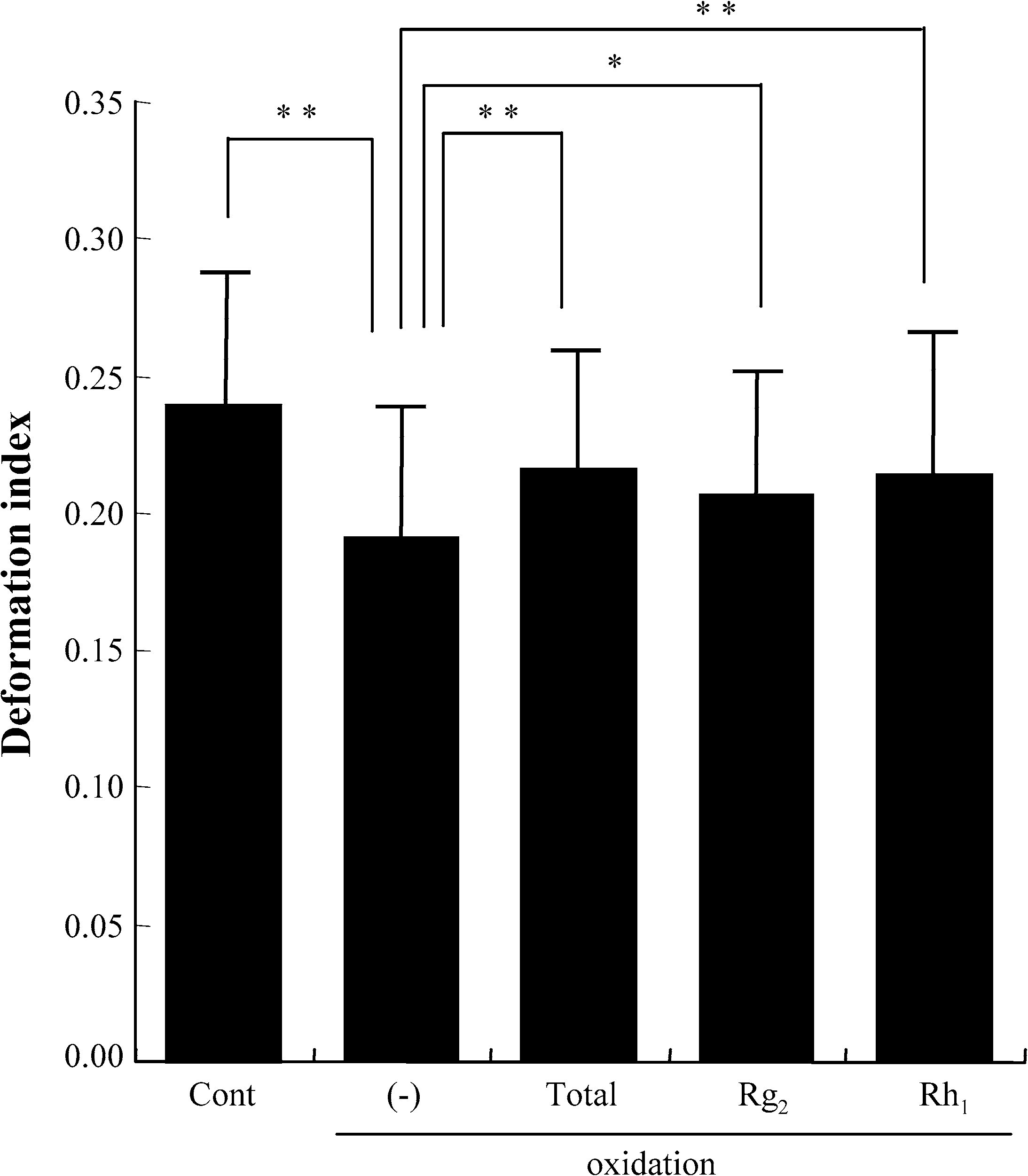 K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 2. Protective effect of Rg2 and Rh1 on oxidation-induced impairment of erythrocyte membrane deformability. Cont: controlunexposed to oxidative stress; total: total ginsenosides; ∗p < 0.05; ∗∗p < 0.01. Values are mean ± SD (n = 10) of thedeformation index.
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 2. Protective effect of Rg2 and Rh1 on oxidation-induced impairment of erythrocyte membrane deformability. Cont: controlunexposed to oxidative stress; total: total ginsenosides; ∗p < 0.05; ∗∗p < 0.01. Values are mean ± SD (n = 10) of thedeformation index.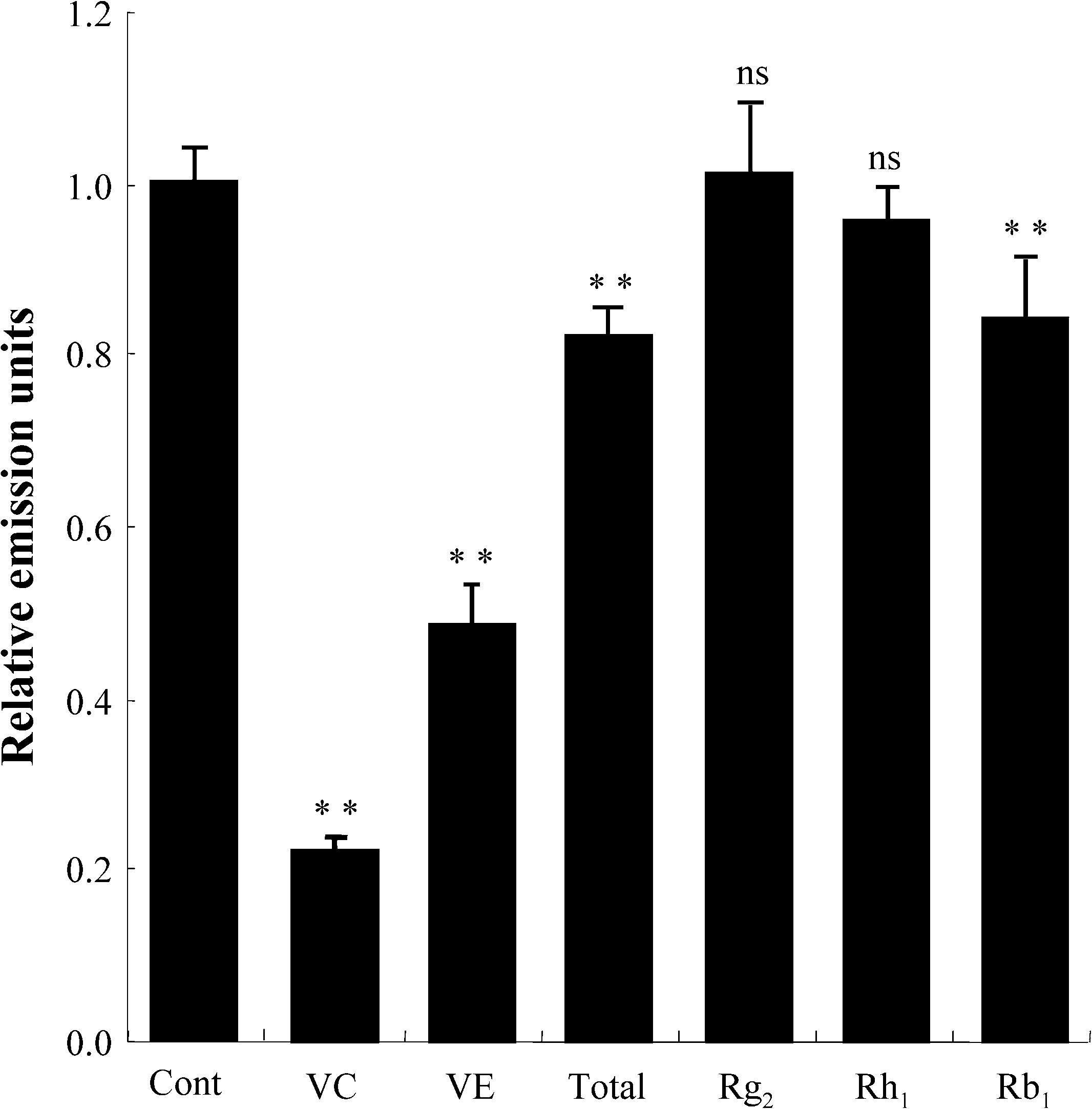
 K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 3. Measurement of antioxidant activity. Data expressed as the ratio of emission units relative to control. The values repre-sented by bars are mean ± SD (n = 20). Cont: control not exposed to oxidative stress; VC: vitamin C; VE: vitamin E; total:total ginsenosides; ∗∗p < 0.01; ns: no significant difference.
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 3. Measurement of antioxidant activity. Data expressed as the ratio of emission units relative to control. The values repre-sented by bars are mean ± SD (n = 20). Cont: control not exposed to oxidative stress; VC: vitamin C; VE: vitamin E; total:total ginsenosides; ∗∗p < 0.01; ns: no significant difference.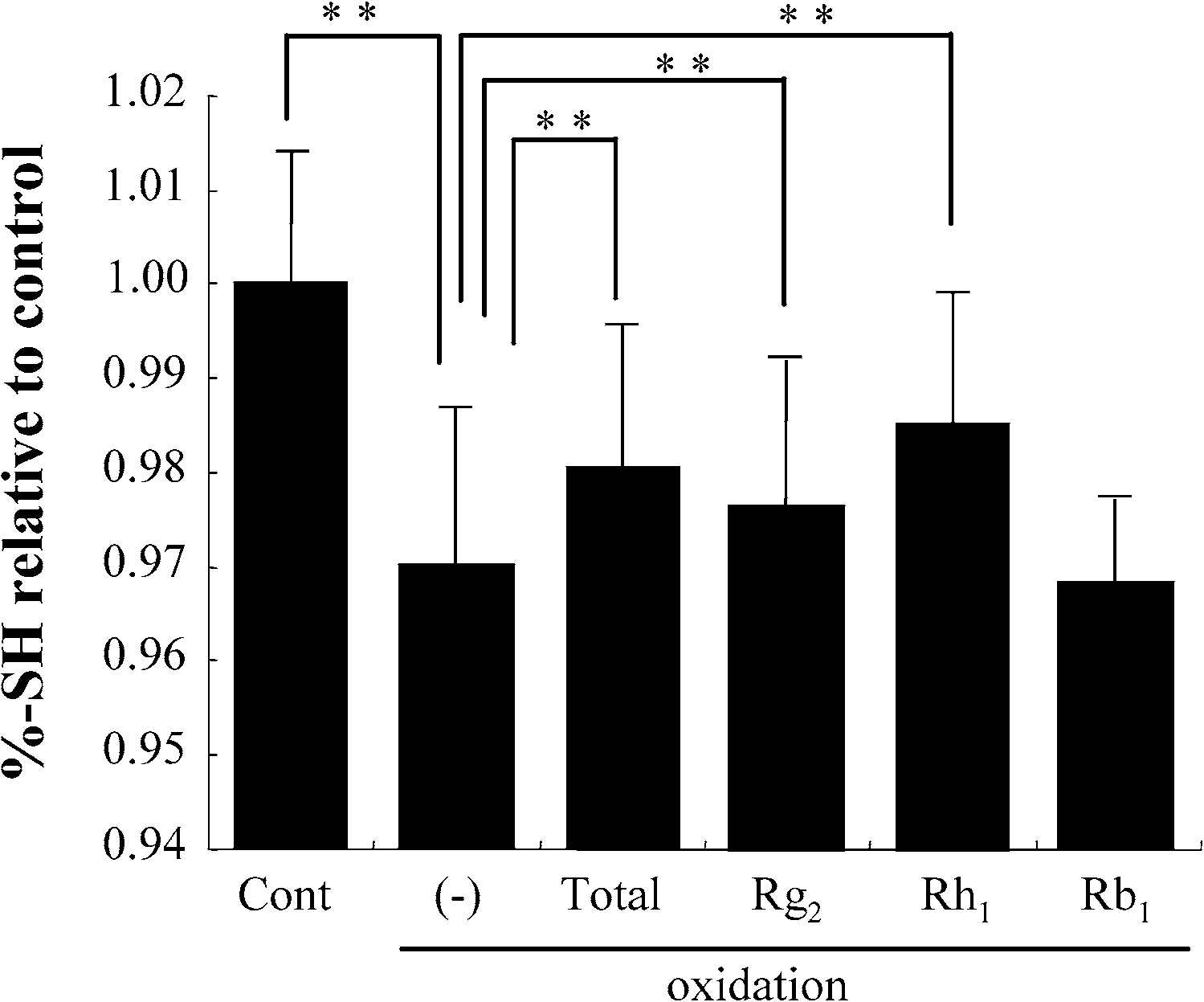
 K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 5. SH-group content in erythrocyte membrane. Cont: control not exposed to oxidative stress; total: total ginsenosides.
K. Samukawa et al. / Ginsenosides Rg2 and Rh1 protect erythrocytes
Fig. 5. SH-group content in erythrocyte membrane. Cont: control not exposed to oxidative stress; total: total ginsenosides.