Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Phyllomedusa-karyotipes.pdf
Italian Journal of Zoology, March 2010; 77(1): 116–121
Karyotypes and Ag-NORs in Phyllomedusa camba De La Riva, 1999and P. rhodei Mertens, 1926 (Anura, Hylidae, Phyllomedusinae):cytotaxonomic considerations
C. R. PAIVA1, J. NASCIMENTO2, A. P. Z. SILVA3, P. S. BERNARDE4, & F. ANANIAS*1
1Curso de Cieˆncias Biolo´gicas, Universidade Sa˜o Francisco (USF), Sa˜o Paulo, Brazil, 2Curso de Cieˆncias Biolo´gicas,Universidade Braz Cubas (UBC), Sa˜o Paulo, Brazil, 3Laborato´rio de Ecologia e Evoluc¸a˜o, Instituto Butantan, Sa˜o Paulo,Sa˜o Paulo, Brazil, and 4Laborato´rio de Herpetologia, Centro de Cieˆncias Biolo´gicas e da Natureza, Universidade Federal doAcre – UFAC, Campus Floresta, Cruzeiro do Sul, Acre, Brazil
AbstractThe karyotypes of Phyllomedusa camba De La Riva, 1999 and P. rhodei Mertens, 1926 are presented and the chromosomepairs with Ag-NORs are identified. Both karyotypes have 2n526 chromosomes with similar morphology, an exceptionbeing the presence of three acrocentric pairs in P. camba. In this species the Ag-NORs are found in the proximal region ofpairs 1 and 5 whilst in P. rhodei an extensive inter-individual variation was observed in the number and position of the Ag-NORs (1p, 3q, 5p, 8p, 11q, and 12q). Based on comparative cytogenetic data of P. camba and P. rhodei, we discuss thephenetic groups proposed for Phyllomedusa genus.
Keywords: Cytogenetic, chromosome, Amphibia, Phyllomedusa, phenetic group
the species can be distributed amongst five speciesgroups: burmeisteri, hypochondrialis, buckeli, perinesos
The family Hylidae has about 870 species, currently
and tarsius (Faivovich et al. 2005; Caramaschi 2006;
distributed in the subfamilies Hylinae, Pelodryadinae
Frost 2007). A few of the Brazilian species were
and Phyllomedusinae (Faivovich et al. 2005; Frost
allocated to other genera such as Hylomanthis,
obtained by Faivovich et al. (2005) show a sister
Phasmahyla and Phrynomedusa by Cruz (1990). The
Downloaded By: [de Pietri, Ana Paula Zampieri S] At: 12:40 12 March 2010
relationship between Pelodryadinae and Phyllome-
phylogenetic relationships between the species in
dusinae, which together correspond to the sister taxon
these groupings are hitherto not well established.
of Hylinae. The subfamily Phyllomedusinae is
Phyllomedusa camba De la Riva, 1999 distributed
comprised of seven nominal genera: Agalychnis,
throughout the southeastern Amazon Basin from
Hylomantis, Cruziohyla, Pachymedusa, Phasmahyla,
Amazon, Acre, and Rondoˆnia) to eastern Bolivia,
Faivovich et al. (2005) discussed several other
is included in the P. tarsius species group together
characters which are likely to be synapomorphies of
with P. boliviana, P. coelestis, P. tarsius, P. trinitatis
Phyllomedusinae, and demonstrated on the basis of
and P. venusta. Barrio-Amoro´s (2006) defined the
molecular data that Cruziohyla is the sister taxon
species belonging to the P. tarsius group using
of the remaining genera. Two clades were observed:
morphological characters (P. coelestis, P. tarsius, P.
one containing Phasmahyla and Phyllomedusa, and the
neildi sp. nov., P. trinitatis, and P. venusta) and
other containing Agalychnis, Hylomantis, Cruziohyla,
proposed that P. boliviana, P. camba and P. sauvagii
Pachymedusa and Phrynomedusa. The 30 species of
(considered in the group by De la Riva 1999, and
Phyllomedusa form a monophyletic group and most of
Faivovich et al. 2005) should be excluded from the
*Correspondence: F. Ananias, Rua Abı´lio Ferraro, 237, 13140 000, Paulı´nia, SP Brazil. Tel: +55 19 3884 7026. Email: feananias@hotmail.com
ISSN 1125-0003 print/ISSN 1748-5851 online # 2010 Unione Zoologica ItalianaDOI: 10.1080/11250000903187585
group because they do not share the most striking
occurrence of a triploid hybrid. In addition, these
feature (herein considered as a synapomorphy) of
authors described the BrdU replication banding
the group, that is, the golden iris with black
reticulations. Although Barrio-Amoro´s (2006) pro-
Considering the difficulty in defining the groups of
posed the exclusion of P. camba from the P. tarsius
Phyllomedusa species as well as the uncertain
species group, its position in the group is maintained
relationships among species of the P. tarsius group,
we present for the first time the karyotypes of P.
Phyllomedusa rohdei Mertens, 1926 is distributed
camba and P. rohdei. Additionally, we also provide
throughout the lowlands of southern Brazil and is
Ag-NOR staining for the two species and observe
included in the P. hypochondrialis species group with
multiple Ag-NORs patterns in the chromosomes of
P. ayeaye, P. azurea, P. centralis, P. hypochondrialis,
P. megacephala, P. nordestina, P. oreades, and P. palliata (Caramaschi 2006; Frost 2007). The speciesP. rhodei has the muscle epicoracoideus, considered
a synapomorphy for the group by Faivovich et al.
Cytogenetic analysis was carried out on two speci-
(2005). Although P. rhodei was included in the study
mens of Phyllomedusa camba collected in Ministro
by Faivovich et al. (2005), the authors did not
Andreazza, state of Roˆndonia (RO) (11u049270 S;
present a phylogenetic relationship between this
61u319010 W) north of Brazil and on seven speci-
species and others in the P. hypochondrialis species
mens of Phyllomedusa rohdei collected in Biritiba-
group, nor with species belonging to other species
Mirim, state of Sa˜o Paulo (SP) (23u349210 S;
46u029190 W), southeastern Brazil (Figures 1A,B).
With regard to cytogenetic data, most of the
The voucher specimens were deposited in the
Amphibian collection (CFBH) of the Departa-
Phyllomedusa is limited to the diploid number, which
is 2n526 for most of the species (Bec¸ak et al. 1970;
Batistic et al. 1975; Batistic 1989; Kuramoto 1990).
The mitotic chromosomes were obtained from
Concerning differential staining technique, Batistic
direct preparations of bone marrow, liver and testis
(1989) presented C-banding and Ag-NOR data and
treated with 0.01% colchicine at a proportion of
suggested a hypothesis for the origin of polyploidy in
0.1 ml/10 g body weight, as described in Baldissera
Phyllomedusa based on the position of the Ag-NORs.
et al. (1993) and Silva et al. (2000), or from
Morando and Hernando (1997) applied the Ag-
NOR technique to chromosomes of P. sauvagii and
(1978). To improve the mitotic index, we injected
P. hypochondrialis and detected differences in the
phytohemagglutinin in some specimens before the
number of Ag-NORs between the species as well as a
colchicine treatment, at the proportion of 0.1 ml/10 g
heteromorphism in P. hypochondrialis. Kasahara et al.
body weight, 48–72 h before sacrifice. Conventional
(2007) demonstrated the occurrence of Ag-NORs in
staining was made with Giemsa 10% diluted in
P. distincta and P. tetraploidea and registered the
phosphate buffer pH 6.8, and silver nitrate labeling
Downloaded By: [de Pietri, Ana Paula Zampieri S] At: 12:40 12 March 2010
Figure 1. A, adult male of Phyllomedusa camba from Ministro Andreazza, Roˆndonia (RO); B, Phyllomedusa rohdei from Biritiba-Mirim, Sa˜oPaulo (SP).
of the nucleolar organizer regions (Ag-NOR) was
As stated by Bogart (1991), centric fusions and
obtained by the technique of Howell and Black
fissions are the most likely mechanism for changes in
chromosomal number. The subfamily Phyllome-dusinae is considered basal in the Hylidae family dueto morphological characters and the diploid number
of 26 chromosomes (Bogart 1973; Morescalchi 1990). Nevertheless, in the species of Phyllomedusa consid-
The species P. camba and P. rhodei have a diploid
ered to be derived, no alterations have occurred in the
number corresponding to 2n526, and a karyotype
number of chromosomes, although the presence of
similar to that found for the majority of the
acrocentric chromosomes and polyploidization in
Phyllomedusinae species: Phyllomedusa (Bec¸ak et
some species suggest the presence of speciation events
al. 1970; Batistic et al. 1975; Batistic 1989;
in this anuran group. The species P. tarsius and P.
Morando & Hernando 1997; Kasahara et al.
camba, both of them with acrocentric chromosomes,
can be considered very closely related and derived in
Pachymedusa (Schmid 1980). An exception was
observed in several tetraploid populations of P.
In P. camba, 20 metaphases showed Ag-NORs in
burmeisteri from Brazil, with 4n552 (Bec¸ak et al.
the proximal regions of the short arms of pair 1, and in
1970; Batistic et al. 1975; Batistic 1989). The
the proximal regions of the long arms of pair 5
karyotype of P. rhodei is made up of metacentric
coincident with secondary constriction (Figure 3B).
pairs (1, 4, 10, and 13), and submetacentric pairs (2,
In P. rohdei, the silver staining showed an extensive
3, 5–9, 11 and 12), while in P. camba, pairs 1, 4, 7,
inter-individual variation in the number and position
10, and 13 are metacentric, 2, 3, 5, 6, and 9 are
of Ag-positive regions, in 1p, 3q, 5p, 8p, 11q and 12q
submetacentric, and 8, 11 and 12 are acrocentric
(Figure 3A). Inter-individual variation in single or
(Figures 2A,B). The occurrence of acrocentric pairs
multiple Ag-NOR patterns was described by Wiley
in P. tarsius observed by Batistic (1989) suggests the
et al. (1989) in Hyla chrysoscelis and H. versicolor,
proximity of this specie and P. camba. Nevertheless,
Foote et al. (1991) in Bufo terrestris, Miura (1994) in
these data refute the proposal of Barrio-Amoro´s
Rana japonica, Schmid et al. (1995) in Agalychnis
(2006), which excluded P. camba from the P. tarsius
callidryas, Kaiser et al. (1996) in Dendropsophus
group. If the presence of acrocentric pairs is
ebraccatus, Silva et al. (1999) in Physalaemus cuvieri,
considered a synapomorphy, P. sauvagii, which
and Silva et al. (2006) in Leptodactylus mystacinus.
possesses no acrocentric chromosomes (Batistic
In Anura, NOR analysis by silver staining has
1989), might be excluded from the P. tarsius group.
shown that species, in both primitive and derived
The karyotype with metacentric and submetacentric
families, possess only one pair of Ag-NORs in their
chromosomes in P. rohdei is compatible to that
found by Batistic (1989) for this species and for P.
Robinson 1986). This observation led King et al.
ayeaye and P. hypochondrialis (all allocated to the P.
(1990) to suggest the presence of only a single pair of
Ag-NORs in diploid karyotypes as an ancestral
Downloaded By: [de Pietri, Ana Paula Zampieri S] At: 12:40 12 March 2010
Figure 2. A, Giemsa-stained karyotypes of Phyllomedusa rohdei; B, Phyllomedusa camba. Note three acrocentric pairs (8, 11 and 12) in P. camba.
Figure 3. A, Metaphases with Ag-NOR bearing chromosomes of Phyllomedusa rohdei; B, Phyllomedusa camba.
condition in Anura, a hypothesis previously pro-
NOR dispersion in anuran genomes may include
posed by Schmid (1978) for bufonids and hylids. In
inversions and translocations involving chromoso-
the genus Phyllomedusa, the occurrence of more than
mal segments containing NORs, transpositions by
one chromosome with Ag-NORs was observed in P.
mobile genetic elements, amplifications of ‘orphan’
burmeisteri, Phyllomedusa sp., P. distincta, P. iheringi,
rDNA cistron, and reinsertion errors during extra-
P. hypochondrialis, P. ayeaye, P. tarsius, P. rohdei and
chromosomal amplifications of ribosomal cistrons.
Incongruous FISH with rDNA probe data with Ag-
Cytochemical tests have revealed that silver does
staining findings were reported in Leptodactylus
not attach to the ribosomal DNA itself, but rather to
mystacinus by Silva et al. (2006), and these authors
proteins specifically associated with this region and
suggested that some Ag-positive sites in the genome
to the heterochromatin (Nardi et al. 1978; Sa´nchez
of L. mystacinus are not true Ag-NORs, but might be
et al. 1995; Dobigny et al. 2002). Nevertheless, in
related to some peculiarities in the heterochromatin
some anuran species, multiple variable Ag-NOR
located at these sites, although they are not always
positive region was confirmed using more suitable
techniques, such as in situ hybridization with a
The present cytogenetic data are not conclusive
fluorescent rDNA probe (Foote et al. 1991; Kaiser
for deciding the taxonomic status of Phyllomedusa at
Downloaded By: [de Pietri, Ana Paula Zampieri S] At: 12:40 12 March 2010
et al. 1996; Lourenc¸o et al. 1998, 2000). These
phenetic species group. Nevertheless, we think that
authors suggest that some mechanisms involved in
our cytogenetic data are toward a closer proximity
Table I. Multiples Ag-NORs sites in Phyllomedusa species.
Batistic 1989; Morand & Hernando 1997
between P. camba and P. tarsius. Cytogenetic studies
Frost DR. 2007. Amphibian species of the world: An online
of other species of the P. tarsius group (sensu De la
reference [Internet]. American Museum of Natural History,New
Riva 1999) are necessary to confirm our hypothesis.
herpetology/amphibia/index.html (accessed 23 July 2007).
Howell WM, Black DA. 1980. Controlled silver-staining of
nucleolus organizer regions with a protective colloidal devel-
oper: A 1-step method. Experientia 36:1014–1015.
Kaiser H, Mais C, Bolanos F, Steinlein C, Feitchtinger W,
Schmid M. 1996. Chromosomal investigation of three Costa
Marcos Yamamoto for collecting the specimens,
Rican frogs from the 30-chromosome radiation of Hyla with
Votorantim Celulose e Papel S/A for providing access
the description of a unique geographic variation in nucleolus
to animals in the Fazenda Sa˜o Sima˜o and Instituto
organizer regions. Genetica 98:95–102.
Brasileiro do Meio Ambiente e Recursos Naturais
Kasahara S, Campos JRC, Catroli GF, Haddad CFB. 2007.
Renova´ veis (IBAMA) for collecting license
Phyllomedusa tetraploidea (2n54x552) and their triploid
(02010.005970/03-69). A.P.Z. Silva acknowledges
FAPESP for financial support (proc. 2006/06634-8).
King M, Contreras N, Honeycutt RL. 1990. Variation within and
between nucleolar regions in Australian hylid frogs (Anura)
shown by 18s and 28s ‘in-situ’ hybridization. Genetica 80:17–29.
Baldissera FA, Oliveira PLS, Kasahara S. 1993. Cytogenetics of
Kuramoto M. 1990. A list of chromosome numbers of anuran
four brazilian Hyla species (Amphibia – Anura) and descrip-
tion of a case with a supernumerary chromosome. Revista
Brasileira de Gene´tica 16:335–345.
Barrio-Amoro´s CL. 2006. A new species of Phyllomedusa (Anura:
Cytogenetics of two species of Paratelmatobius (Anura:
Hylidae: Phyllomedusinae) from northweastern Venezuela.
Leptodactylidae), with phylogenetic comments. Hereditas
Batistic RF. 1989. Aspectos citogene´ticos da evoluc¸a˜o em
Ribeira˜o Preto (Brazil): Insyituto Butantan. pp 203.
Batistic RF, Soma M, Bec¸ak ML, Bec¸ak W. 1975. Further studies
detected by silver staining and fluorescence in situ hybridiza-
tion. Chromosome Research 6:621–628.
Phyllomedusa burmeisteri. Journal of Heredity 66:160–162.
Mahony MJ, Robinson ES. 1986. Nucleolar organiser region
Bec¸ak ML, Denaro L, Bec¸ak W. 1970. Polyploidy and mechan-
(NOR) location in karyotypes of Australian ground frogs
isms of karyotypic diversification in Amphibia. Cytogenetics
(Family Myobatrachidae). Genetica 68:119–127.
Miura I. 1994. Sex chromosome differentiation in the Japanese
Bogart JP. 1973. Evolution of anuran karyotypes. In: Vial JL,
Brown frog, Rana japonica. II. Sex-linkage analyses of the
editor. Evolutionary biology of anurans. Columbia: University
nucleolar organizer regions in chromosomes no. 4 of the
Bogart JP. 1991. The influence of life history on karyotypic
evolution in frogs. In: Green DM, Sessions SK, editors.
Amphibian cytogenetics and evolution. San Diego, CA:
genes ribosomales activos en Phyllomedusa hypochondrialis y P.
sauvagii (Anura: Hylidae). Cuadernos Herpetologia 11:31–36.
Caramaschi U. 2006. Redefinic¸a˜o do grupo de Phyllomedusa
Morescalchi A. 1990. Cytogenetics and the problem of lissam-
hypocondrialis, com redescric¸a˜o de P. megacephala (Miranda-
phibian relationships. In: Olmo E, editor. Cytogenetics of
Downloaded By: [de Pietri, Ana Paula Zampieri S] At: 12:40 12 March 2010
Ribeiro, 1926), revalidac¸a˜o de P. azurea Cope, 1862 e
amphibians and reptiles. Birkha¨ser: Verlag. pp 1–19.
descric¸a˜o de uma nova espe´cie (Amphibia, Anura, Hylidae).
Nardi I, de Lucchini S, Barsacchi-Pilone G, Andronico F. 1978.
Arquivos do Museu Nacional do Rio de Janeiro 64:159–179.
Chromosome location of the ribosomal RNA genes in Triturus
Phyllomedusinae da Floresta Atlaˆntica (Amphibia, Anura,
Hylidae). Revista Brasileira de Biologia 50:709–726.
Sa´nchez A, Jimenez R, Burgos M, Stitou S, Zurita F, de la
De la Riva I. 1999. A new Phyllomedusa from southweastern
Guardia RD. 1995. Cytogenetic peculiarities in the Algerian
Amazonia (Amphibia, Anura: Hylidae). Revista Espan
hedgehog: Silver stains not only NORs but also heterochro-
Dobigny G, Ozouf-Costaz C, Bonillo C, Voloubouev V. 2002.
Schmid M. 1978. Chromosome banding in Amphibia. I.
‘Ag-NORs’ are not always true NORs: New evidence in
Constitutive heterochromatin and nucleolus organizers regions
mammals. Cytogenetic and Genome Research 98:75–77.
in Bufo and Hyla. Chromosoma 66:361–388.
Faivovich J, Haddad CFB, Garcia PCA, Frost D, Campbell JA,
Schmid M. 1980. Chromosome banding in Amphibia. IV.
Wheeler WC. 2005. Systematic review of the frog family
Differentiation of GC- and AT-rich chromosome regions in
Hylidae, with special reference to Hylinae: Phylogenetic
analysis and taxonomic revision. Bulletin of the American
Schmid M. 1982. Chromosome banding in Amphibia VII.
Museum of Natural History 294:1–240.
Analysis of the structure and variability of NORs in Anura.
Foote DL, Wiley JE, Little ML, Meyne J. 1991. Ribosomal RNA
genes site polymorphism in Bufo terrestris. Cytogenetic Cell
Schmid M, Feichtinger W, Wimer R, Mais C, Bolan
1995. Chromosome banding in Amphibia XXI. Inversion
polymorphism and nucleolus organizer regions in Agalychnis
evidence of a unique case of Ag-NOR variability. Hereditas
callidryas (Anura, Leptodactylidae). Chromosoma 101:284–292.
Silva APZ, Haddad CFB, Galassi GG, Kasahara S. 2006.
Silva APZ, Haddad CFB, Kasahara S. 2000. Chromosomal studies
Multiple nucleolus organizer regions in Leptodactylus mystaci-
on five species of the genus Leptodactylus Fitzinger, 1826 (Amphi-
nus (Amphibia, Anura) and comments on its systematic
bia, Anura) using differential staining. Cytobios 103:25–38.
position in the L. fuscus group based on cytogenetic and
Wiley JE, Little ML, Romano MA, Blount DA, Cline GR. 1989.
molecular analyses. Genetica 127:35–44.
Polymorphism in the location of the 18S and 28S rDNA genes
Silva APZ, Haddad CFB, Kasahara S. 1999. Nucleolus organizer
on the chromosomes of the diploid–tetraploid treefrogs Hyla
regions in Physalaemus cuvieri (Anura, Leptodactylidae), with
chrysoscelis and Hyla versicolor. Chromosoma 97:481–487.
Downloaded By: [de Pietri, Ana Paula Zampieri S] At: 12:40 12 March 2010
EdxTM Metodi di diagnostica e trattamento energetici Capitolo tratto da “Energy Psychology in Psychotherapy”, 2002, W.W.Norton di Fred Gallo Nel 1992, dopo aver lavorato per 22 anni come psicoterapeuta ho conosciuto tre approcci che hanno influenzato profondamente il mio modo di pensare e di fare terapia oggi: l’EMDR (Desensibilizzazione e Rielaborazione attraverso i Movimenti Ocular
Written by Ian Maber Educational Consultant for PolyVision EMEA (Europe Middle East & Africa) June 2009 • Introducing the IWB (again!) In a world of technological advancements the adoption of IWB’s into the classroom is becoming an integral part of the everyday work of an educationalist/teacher. Advisors and consultants who fully understand the education market will always agre
 Italian Journal of Zoology, March 2010; 77(1): 116–121
Karyotypes and Ag-NORs in Phyllomedusa camba De La Riva, 1999and P. rhodei Mertens, 1926 (Anura, Hylidae, Phyllomedusinae):cytotaxonomic considerations
C. R. PAIVA1, J. NASCIMENTO2, A. P. Z. SILVA3, P. S. BERNARDE4, & F. ANANIAS*1
1Curso de Cieˆncias Biolo´gicas, Universidade Sa˜o Francisco (USF), Sa˜o Paulo, Brazil, 2Curso de Cieˆncias Biolo´gicas,Universidade Braz Cubas (UBC), Sa˜o Paulo, Brazil, 3Laborato´rio de Ecologia e Evoluc¸a˜o, Instituto Butantan, Sa˜o Paulo,Sa˜o Paulo, Brazil, and 4Laborato´rio de Herpetologia, Centro de Cieˆncias Biolo´gicas e da Natureza, Universidade Federal doAcre – UFAC, Campus Floresta, Cruzeiro do Sul, Acre, Brazil
AbstractThe karyotypes of Phyllomedusa camba De La Riva, 1999 and P. rhodei Mertens, 1926 are presented and the chromosomepairs with Ag-NORs are identified. Both karyotypes have 2n526 chromosomes with similar morphology, an exceptionbeing the presence of three acrocentric pairs in P. camba. In this species the Ag-NORs are found in the proximal region ofpairs 1 and 5 whilst in P. rhodei an extensive inter-individual variation was observed in the number and position of the Ag-NORs (1p, 3q, 5p, 8p, 11q, and 12q). Based on comparative cytogenetic data of P. camba and P. rhodei, we discuss thephenetic groups proposed for Phyllomedusa genus.
Italian Journal of Zoology, March 2010; 77(1): 116–121
Karyotypes and Ag-NORs in Phyllomedusa camba De La Riva, 1999and P. rhodei Mertens, 1926 (Anura, Hylidae, Phyllomedusinae):cytotaxonomic considerations
C. R. PAIVA1, J. NASCIMENTO2, A. P. Z. SILVA3, P. S. BERNARDE4, & F. ANANIAS*1
1Curso de Cieˆncias Biolo´gicas, Universidade Sa˜o Francisco (USF), Sa˜o Paulo, Brazil, 2Curso de Cieˆncias Biolo´gicas,Universidade Braz Cubas (UBC), Sa˜o Paulo, Brazil, 3Laborato´rio de Ecologia e Evoluc¸a˜o, Instituto Butantan, Sa˜o Paulo,Sa˜o Paulo, Brazil, and 4Laborato´rio de Herpetologia, Centro de Cieˆncias Biolo´gicas e da Natureza, Universidade Federal doAcre – UFAC, Campus Floresta, Cruzeiro do Sul, Acre, Brazil
AbstractThe karyotypes of Phyllomedusa camba De La Riva, 1999 and P. rhodei Mertens, 1926 are presented and the chromosomepairs with Ag-NORs are identified. Both karyotypes have 2n526 chromosomes with similar morphology, an exceptionbeing the presence of three acrocentric pairs in P. camba. In this species the Ag-NORs are found in the proximal region ofpairs 1 and 5 whilst in P. rhodei an extensive inter-individual variation was observed in the number and position of the Ag-NORs (1p, 3q, 5p, 8p, 11q, and 12q). Based on comparative cytogenetic data of P. camba and P. rhodei, we discuss thephenetic groups proposed for Phyllomedusa genus. group because they do not share the most striking
occurrence of a triploid hybrid. In addition, these
feature (herein considered as a synapomorphy) of
authors described the BrdU replication banding
the group, that is, the golden iris with black
reticulations. Although Barrio-Amoro´s (2006) pro-
Considering the difficulty in defining the groups of
posed the exclusion of P. camba from the P. tarsius
Phyllomedusa species as well as the uncertain
species group, its position in the group is maintained
relationships among species of the P. tarsius group,
we present for the first time the karyotypes of P.
group because they do not share the most striking
occurrence of a triploid hybrid. In addition, these
feature (herein considered as a synapomorphy) of
authors described the BrdU replication banding
the group, that is, the golden iris with black
reticulations. Although Barrio-Amoro´s (2006) pro-
Considering the difficulty in defining the groups of
posed the exclusion of P. camba from the P. tarsius
Phyllomedusa species as well as the uncertain
species group, its position in the group is maintained
relationships among species of the P. tarsius group,
we present for the first time the karyotypes of P.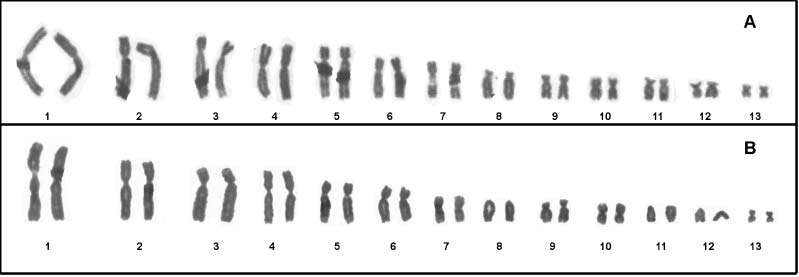 of the nucleolar organizer regions (Ag-NOR) was
As stated by Bogart (1991), centric fusions and
obtained by the technique of Howell and Black
fissions are the most likely mechanism for changes in
chromosomal number. The subfamily Phyllome-dusinae is considered basal in the Hylidae family dueto morphological characters and the diploid number
of 26 chromosomes (Bogart 1973; Morescalchi 1990).
of the nucleolar organizer regions (Ag-NOR) was
As stated by Bogart (1991), centric fusions and
obtained by the technique of Howell and Black
fissions are the most likely mechanism for changes in
chromosomal number. The subfamily Phyllome-dusinae is considered basal in the Hylidae family dueto morphological characters and the diploid number
of 26 chromosomes (Bogart 1973; Morescalchi 1990).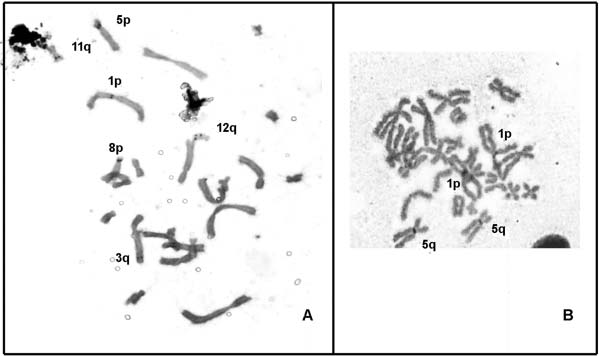 Figure 3. A, Metaphases with Ag-NOR bearing chromosomes of Phyllomedusa rohdei; B, Phyllomedusa camba.
Figure 3. A, Metaphases with Ag-NOR bearing chromosomes of Phyllomedusa rohdei; B, Phyllomedusa camba.