Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Hepatitiscentral.net
Ting Zhang,1,2 Yuan Li,1 Jian-Ping Lai,1 Steven D. Douglas,1 David S. Metzger,3 Charles P. O’Brien,3 and Wen-Zhe Ho1
Alcohol consumption accelerates liver damage and diminishes the anti– hepatitis C virus (HCV) effect of interferon alfa (IFN-␣) in patients with HCV infection. It is unknown, however, whether alcohol enhances HCV replication and promotes HCV disease progres- sion. The availability of the HCV replicon containing hepatic cells has provided a unique opportunity to investigate the interaction between alcohol and HCV replicon expression. We determined whether alcohol enhances HCV RNA expression in the replicon containing hepatic cells. Alcohol, in a concentration-dependent fashion, significantly increased HCV replicon expression. Alcohol also compromised the anti-HCV effect of IFN-␣. Investigation of the mechanism(s) responsible for the alcohol action on HCV replicon indicated that alcohol activated nuclear factor B (NF-B) promoter. Caffeic acid phenethyl ester (CAPE), a specific inhibitor of the activation of NF-B, abolished alcohol-induced HCV RNA ex- pression. In addition, naltrexone, an opiate receptor antagonist, abrogated the enhancing effect of alcohol on HCV replicon expression. In conclusion, alcohol, probably through the activation of NF-B and the endogenous opioid system, enhances HCV replicon expression and compromises the anti-HCV effect of IFN-␣. Thus, alcohol may play an important role in vivo as a cofactor in HCV disease progression and compromise IFN-␣– based therapy against HCV infection. (HEPATOLOGY 2003;38:57-65.)
HepatitisCvirus(HCV)isresponsibleforthe cipientsoflivertransplantation.HCV,firstmolecularly
vast majority of cases of transfusion-associated
cloned in 1989,1 is a positive-strand RNA virus of the
and community-acquired non-A, non-B hepa-
flavivirus family with a genome size of ϳ10 kb, which
titis1,2 and infects an estimated 170 million people world-
encodes a number of structural (core, E1, E2, and p7) and
wide.3 The seroprevalence of anti-HCV antibody in the
nonstructural (NS2, NS3, NS4A, NS4B, NS5A and
United States has been estimated at 1.8%, which corre-
NS5B) proteins.6 HCV has at least 6 distinct but related
sponds to approximately 4 million people.4 HCV is the
genotypes with more than 50 subtypes, and genotype 1 is
leading cause of chronic viral hepatitis in the United
the most common in the United States, Europe, and most
States,5 and HCV-infected individuals are the major re-
parts of Asia. HCV typically escapes clearance by thehost’s immune system and leads to the establishment of apersistent infection in approximately 70% of infected in-
Abbreviations: HCV, hepatitis C virus; IFN-␣, interferon alfa; CAPE, caffeic
dividuals.7 The consequences of a subset of patients with
acid phenethyl ester; 4-MP, 4-methylpyrazole; IgG, immunoglobulin G; RT-PCR,
chronic HCV infection are cirrhosis, liver failure, and
reverse-transcription polymerase chain reaction; mRNA, messenger RNA; NF-B,nuclear factor kappa B; 5Ј-NCR, 5Ј-noncoding region; MB, molecular beacon.
hepatocellular carcinoma.8,9 Treatment of HCV with in-
From the 1Division of Allergy and Immunology, Joseph Stokes Jr. Research Insti-
terferon alfa (IFN-␣) and ribavirin is associated with a
tute at The Children’s Hospital of Philadelphia, Department of Pediatrics, Univer-
sustained response rate of less than 50%.7,10,11 These lim-
sity of Pennsylvania School of Medicine, Philadelphia, PA; the 2Departmentof Infectious Diseases, The Children’s Hospital of Fudan University, Shanghai,
ited therapeutic efficacies and the absence of an effective
China; and the 3Department of Psychiatry, The Center for Studies of Addiction,
HCV vaccine underscore the importance of research on
University of Pennsylvania School of Medicine, Philadelphia, PA.
factors that enhance HCV infection and compromise
Received November 20, 2002; accepted April 24, 2003.Supported by grants from the National Institutes of Health (DA12815 andDA16022 to W.-Z. Ho; MH49981 and AA13547 to S.D. Douglas).
Alcohol is the most commonly used and abused drug
Address reprint requests to: Dr. Wen-Zhe Ho, Division of Allergy and Immunol-
in the United States. Alcohol abuse significantly affects
ogy, The Children’s Hospital of Philadelphia, University of Pennsylvania School ofMedicine, 34th St. and Civic Center Blvd., Philadelphia, PA 19104. E-mail:
morbidity and mortality from infectious diseases.12 Alco-
ho@email.chop.edu; fax: 215-590-2025.
hol consumption accelerates liver damage, diminishes
Copyright 2003 by the American Association for the Study of Liver Diseases.
hepatocellular carcinoma in patients with chronic HCV
infection.13-18 Alcohol added in vivo and in vitro also im-
Southern blots are negative for replicon nucleic acid inte-
pairs liver parenchymal cells and various functions of im-
grated into the host chromosome in Huh.8 cells.21 Using
mune cells, including monocytes, T cells, and natural
a real-time reverse-transcription polymerase chain reac-
killer cells, which contribute to hepatocyte damage in
tion (RT-PCR) assay that we have recently developed,22
chronic HCV infection.12,19 Alcohol consumption and
we were able to detect ϳ2,500 to 5,000 copies of HCV
viral hepatitis infection, both recognized as major causes
messenger RNA (mRNA) per Huh.8 cell. Huh.2 is a cell
of liver disease worldwide, frequently coexist in patients
clone containing a G418 selectable HCV replicon with
with chronic liver disease.13,17,18 Alcohol and HCV most
HCV NS5A mutation (S1172P).21 FCA-1 cells contain a
likely act synergistically to promote the development and
subgenomic replicon from a known infectious HCV
progression of liver damage.17,18 There is little direct in-
clone with several consensus mutations in NS3 as well as
formation available concerning the effects of alcohol
in NS5A (NS3: E177G; NS5A: D1229E, I1299V).20
abuse on HCV replication in hepatic cells. This lack of
The HCV replicon containing cells and Huh7 cells were
knowledge about the impact of alcohol abuse on HCV is
maintained in Dulbecco Modified Eagle’s Medium
a major barrier to fundamental understanding of HCV-
(Gibco-BRL, Grand Island, NY) supplemented with
related morbidity and mortality in alcohol abusers with
10% (vol/vol) fetal bovine serum (HyClone, Logan, UT),
HCV infection. Thus, it is critical to investigate the im-
100 U/mL penicillin, 100 g/mL streptomycin, 2
pact of alcohol abuse on HCV replication in the target
mmol/L L-glutamine, and 0.1 mmol/L minimum essen-
cells such as hepatic cells. We investigated whether alco-
tial medium nonessential amino acids in a humidified
hol enhances HCV RNA expression in HCV replicon
containing cell lines.20,21 We also studied whether the in
medium for Huh.8, Huh.2, and FCA-1 cells contained
vitro addition of alcohol to these cells compromises the
G418 (750 g/mL). Cells were passaged every 2 or 3 days
and seeded in 24-well plates at a density of 105 cells/well. Alcohol Treatment. The HCV replicon containing Materials and Methods
cells (Huh.8, Huh.2, and FCA-1) and Huh7 cells platedin 24-well plates were incubated with or without alcohol
Reagents. Alcohol was purchased from Aaper Alcohol
(ϳ10 to 100 mmol/L) for up to 96 hours. For multiple
and Chemical Company (Shelbyville, KY). Recombinant
time-point treatment, the cells were treated with alcohol
IFN-␣ was obtained from R&D Systems Inc. (Minneap-
daily. To study whether alcohol interferes with the anti-
olis, MN). The opiate receptor antagonist naltrexone was
HCV effect of IFN-␣, Huh.8 cells were incubated with
obtained from Sigma (St. Louis, MO). Caffeic acid phen-
alcohol (100 mmol/L) and/or IFN-␣ (100 U/mL).
ethyl ester (CAPE) was purchased from Calbiochem-No-
IFN-␣ was added to the cell cultures for 30 minutes be-
vabiochem Corp. (San Diego, CA). 4-Methylpyrazole (4-
fore the addition of alcohol. We selected IFN-␣ at the
MP) and cyanamide were obtained from Sigma. Mouse
concentration that was tested to have an anti-HCV effect
monoclonal anti-HCV NS5 antibody was a gift obtainedfrom Dr. Bill Sun (Thomas Jefferson University, Phila-
in HCV replicon containing cells.20,21 In the experiments
delphia, PA). Horseradish peroxidase-conjugated goat
to investigate whether CAPE, a specific inhibitor of the
anti-mouse immunoglobulin G (IgG) was purchased
activation of nuclear factor B (NF-B), suppresses the
from Jackson Immune Research Labs (West Grove, PA)
enhancing effect of alcohol on HCV RNA expression,
Cell Lines. Huh.8 and Huh7 cell lines were obtained
Huh.8 cells were incubated with CAPE for 30 minutes
from Dr. Charles Rice (The Rockefeller University, New
before the addition of alcohol. The selected concentration
York, NY, and Apath, L.L.C., St. Louis, MO). Huh.2
of CAPE (20 g/mL) was based on our pilot experiments,
cells were obtained from the National Institutes of Health
which showed that CAPE at the concentrations of 20 to
AIDS Research and Reference Reagent Program. FCA-1
30 g/mL resulted in a maximum inhibitory effect on
cells were obtained from Dr. Christoph Seeger (Fox
HCV RNA expression without cytotoxic effect (trypan
Chase Cancer Center, Philadelphia, PA). Huh7, a human
blue dye staining) on Huh.8 cells (data not shown). In
hepatoma cell line, is the parental cell line of Huh.8,
addition, our pilot experiments showed that there was no
Huh.2, and FCA-1. Huh.8 is a cell clone containing a
cytotoxic effect of alcohol at the concentrations of 100
G418 selectable HCV RNA replicon with wild-type
mmol/L23 or less on the HCV replicon containing cells as
HCV nonstructural protein NS5A sequence.21 The rep-
shown by trypan blue dye staining. In some experiments,
licon in Huh.8 cells contains both the 5Ј nontranslating
Huh.8 cells were preincubated with either 4-MP (0.1
region and 3Ј nontranslating region as well as the open
mol/L) or cyanamide (0.2 mol/L) or naltrexone (10Ϫ8
reading frame of the nonstructural proteins NS3-5B.21
mol/L) for 30 minutes before the addition of alcohol.
To minimize alcohol evaporation that diminishes alco-
The PCR buffer contained 5-carboxy-X-rhodamine (500
hol concentration in the plates, we maintained alcohol-
nmol/L) as the reference dye for normalization of the reac-
treated cells in the plates sealed with PARAFILM
tions. Any possible fluctuations in 5-carboxy-X-rhodamine
(American National Can, Greenwich, CT). Furthermore,
signal are used to correct the sample signal. All amplification
to avoid evaporated alcohol contamination of control cul-
reactions were performed in duplicate. A standard curve was
ture plates, alcohol-treated and control plates were main-
generated with 10-fold dilutions of HCV 5Ј-NCR RNA
control22 prequantitated by a spectrophotometer (Eppen-
RNA Extraction. Total RNA (1 g) was extracted
dorf Scientific, Inc., Westbury, NY).
from HCV replicon containing cells as well as from Huh7
Immunoblot Assay. Total cell lysates were prepared
cells using Tri-Reagent (Molecular Research Center, Cin-
from Huh.8 cells (105 cells per well in a 24-well plate)
cinnati, OH). In brief, total RNA was extracted by a
incubated with or without alcohol for 72 hours using a
single-step guanidinium thiocyanate-phenol-chloroform
lysis buffer (Promega, Madison, WI). The protein con-
extraction. After centrifugation at 13,000g for 15 min-
centration was determined by DC protein assay kit (Bio-
utes, the RNA-containing aqueous phase was precipitated
Rad, Hercules, CA). The immunoblot analysis of HCV
in same volume isopropanol. RNA precipitates were then
NS5 protein was performed using a Bio-Dot SF apparatus
washed once in 75% alcohol and resuspended in 20 L of
as described by the manufacturer. Briefly, total protein
(0.5 g) extracted from Huh.8 cells incubated with or
Real-Time RT-PCR for HCV RNA Quantification.
without alcohol (100 mmol/L) was applied onto a nitro-
We used the real-time RT-PCR assay newly developed in
cellulose membrane. After the blockage with phosphate
our laboratory for the quantification of HCV RNA.22 The
buffered saline containing 5% nonfat dry milk for 1 hour
real-time PCR was performed with 1/10th of cDNA de-
at room temperature, the membrane was incubated with a
rived from RNA (1 g) extracted from the cells using ABI
mouse monoclonal anti-HCV NS5 antibody at 4°C over-
Prism 7700 Sequence Detection System (Applied Biosys-
night. After washing 3 times with Tween phosphate-buff-
tems, Foster City, CA). We designed the primer pair
ered saline, the nitrocellulose membrane was incubated
(S130/AS311) that is specific for the recognition of highly
with horseradish peroxidase-conjugated goat anti-mouse
conservative 5Ј-noncoding region (5Ј-NCR) in the HCV
IgG for 1 hour. Bound antibody was visualized by devel-
genome.22 Their sequences are as follows: 5Ј-CGGGAG-
oping the membrane in SuperSignal West Pico chemilu-
AGCCATAGTGGTCTGCG-3Ј (S130) and 5Ј-CTC-
minescent substrate kit (Pierce, Rockford, IL). The results
were recorded on film (Eastman Kodak, Rochester, NY).
probe (molecular beacon [MB]) sequence for HCV 5Ј-NCR
NF-B Promoter Activation Assay. The plasmid
was selected within the primer pair of S130 and AS311,
(pNF-B-Luc) containing NF-B promoter linked with
which was designed to be perfectly complementary to the
a luciferase gene was generated by Dr. Daniel Petrak.24
target sequence in 5Ј-NCR of HCV genome.22 The follow-
Two copies of the mouse light chain enhancer25 were
ing is the sequence of the MB: 5Ј-FAM- GCGAGCCAC-
cloned into pBLCAT3 vector,26 and then the construct
CGGAATTGCCAGGACGACCGCTCGC-DABCYL-3Ј. was modified by replacing the chloramphenicol acetyl-The stem sequence (underlined) of MB does not comple-
transferase reporter with the luciferase gene obtained
ment the sequences within the loop region. The MB was
from pGEM-Luc.24 Plasmid DNA was prepared by Mini-
labeled at the 5Ј end with 6-carboxyfluorescein (6-FAM) and
prep techniques, according to the manufacturer’s instruc-
the quencher 4-(4Ј-dimethylaminophenylaso) benzoic acid
tions (Wizard Plus Minipreps, Promega, Madison, MI)
(DABCYL) at the 3Ј end. Both primers and MB were sus-
and used in transfection experiments. For transfection
pended in Tris-EDTA buffer and stored at Ϫ30°C. Thermal
experiments, Huh.8 and Huh7 cells were seeded in a
cycling conditions were designed as follows: initial denatur-
6-well tissue culture plate at the density of 3 ϫ 105 cells
ation at 95°C for 10 minutes, followed by 40 cycles of 95°C
per well 1 day before the transfection. The cells were
for 15 seconds and 60°C for 60 seconds. Fluorescence mea-
transfected with the pNF-B-Luc using FuGENE 6
surements were recorded during each annealing step. At the
Transfection Reagent (Roche Diagnostic Corporation,
termination of each PCR run, the data were automatically
Indianapolis, IN) with a ratio of FuGENE 6: plasmid 6:1
analyzed by the system and amplification plots were gener-
(L:g). Six hours after the transient transfection, the
ated. For each PCR reaction, 2 L of cDNA template was
cells were incubated with or without alcohol (100
added to 48 L of PCR master mixture (5 L of 1ϫ PCR
mmol/L) for 24 hours. At the termination of the experi-
buffer II, 5 mmol/L MgCl2, 300 nmol/L dNTP, 400
ments, the cells were harvested and washed twice with
nmol/L of each primer, 1.5 U of AmpliTaq Gold DNA
phosphate buffered saline by centrifugation at 3,300g for
polymerase, 400 nmol/L of MBs, and 24.7 L of water).
3 minutes at room temperature. The cell pellets were lysed
Effect of Alcohol on HCV Replicon. To evaluate the
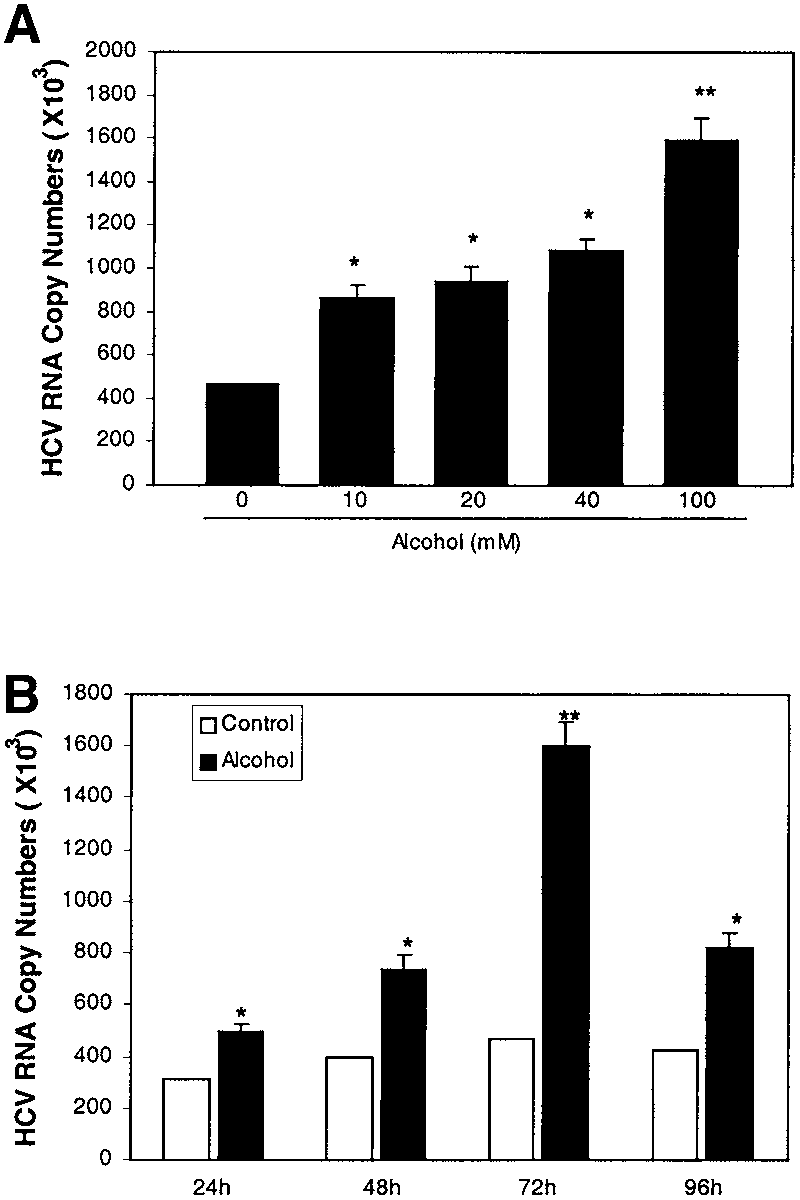
effect of alcohol on HCV RNA expression in HCV rep-licon containing cells, we incubated Huh.8, Huh.2, andFCA-1 cells with or without alcohol. The alcohol concen-trations selected for the study were based on our pilotexperiments, which showed that alcohol at the concentra-tion of 100 mmol/L or lower had no cytotoxic effect onHuh.8 cells (data not shown). Total cellular RNA ex-tracted from the cell cultures was subjected to the real-time RT-PCR for the quantification of HCV RNA levels72 hours after alcohol treatment. The addition of alcoholto Huh.8 cell cultures increased HCV RNA expression ina concentration-dependent fashion (Fig. 1A). To deter-mine whether the effect of alcohol on HCV RNA expres-sion is time dependent, we incubated Huh.8 cells withalcohol (100 mmol/L) and harvested cells at 4 differenttime points (24 hours, 48 hours, 72 hours, and 96 hours)after alcohol treatment for HCV RNA quantification. The maximum HCV RNA expression in alcohol-treatedHuh.8 cells was observed at 72 hours post-treatment (Fig. 1B). Because NS5 protein plays a critical role in HCV
Fig. 1. Effect of alcohol on HCV RNA expression in Huh.8 cells. (A)
replication,21,27 we also examined whether alcohol alters
Effect of alcohol on HCV RNA expression in Huh.8 cells at different
HCV NS5 protein expression in Huh.8 cells using the
concentrations. Huh.8 cells (105 cells/mL/well) plated in a 24-well platewere incubated with or without alcohol at indicated concentrations for 72
immunoblot assay. In comparison with untreated Huh.8
hours. (B) Effect of alcohol on HCV RNA expression in Huh.8 cells at
cells, alcohol (100 mmol/L)-treated cells express higher
different time points. Huh.8 cells were incubated with or without alcohol
levels of NS5 protein as shown by the enhanced intensity
(100 mmol/L). Total cellular RNA extracted from the cell cultures was
of protein band (Fig. 2). To determine whether the en-
subjected to real-time RT-PCR for HCV and glyceraldehyde-3-phosphatedehydrogenase (GAPDH) RNA quantification. The data are expressed as
hancing effect of alcohol on HCV replicon is not cell
HCV RNA copy numbers per reaction, which was normalized based on the
clone dependent, we examined the effect of alcohol on
ratio of HCV RNA/GAPDH mRNA that was amplified in the same plate by
HCV replicon in 2 additional HCV replicon containing
the real-time RT-PCR. The data shown are the mean Ϯ SD of triplicatecultures, and the experiment was repeated 5 times with similar results
cell clones (Huh.2 and FCA-1). Similar to the findings
(*P Ͻ .05; **P Ͻ .01).
observed in Huh.8 cells, alcohol increased HCV RNAexpression in both Huh.2 and FCA-1 cells (Fig. 3). Alcohol Inhibits the Anti-HCV Effect of IFN-␣.
with 0.25 mL of 1ϫ Reporter Lysis Buffer (Promega) and
Alcohol consumption impairs the therapeutic response to
a cycle of freezing and thawing in dry ice. Cell-free lysates
IFN-␣ in patients with HCV infection.14-16 Because
were obtained by centrifugation at 10,000g for 30 secondsat room temperature. The effects of alcohol on the acti-vation of NF-B promoter in these transiently transfectedcells were determined by NF-B promoter-driven lucif-erase activity. Luciferase activity in cell lysate (50 L persample) was quantitated using a Luciferase Assay System(Promega) and a luminometer. The results are presentedas relative light units. Statistical Analysis. All variables were tested in trip-
Fig. 2. Effect of alcohol on HCV NS5 protein expression in Huh.8 cells.
Huh.8 cells (105 cells/mL/well) plated in a 24-well plate were incubated
licate, and experiments were repeated at least 3 times.
with or without alcohol (100 mmol/L) for 72 hours. Cell lysates were
Triplicate wells had variability of less than 15%. One-way
quantified by DC protein assay kit. Equal amounts (0.5 g) of protein
ANOVA was used to test for the difference in means and
extracted from alcohol-treated and untreated (control) Huh.8 cells wereapplied onto a nitrocellulose membrane for immunoblot assay. The
post-hoc t test was used for comparisons. The differences
results were recorded on the film (2-minute exposure). One representa-
were considered significant if P Ͻ .05.
Fig. 3. Effect of alcohol on HCV RNA expression in different HCV
replicon cell clones. Huh.8, Huh.2, and FCA-1 cells (105 cells/mL/well)plated in a 24-well plate were incubated with or without alcohol (100mmol/L) for 72 hours. Total cellular RNA extracted from the cell cultureswas subjected to real-time RT-PCR for HCV and GAPDH RNA quantifica-tion. The data are expressed as HCV RNA copy numbers per reaction,which was normalized based on the ratio of HCV RNA/GAPDH mRNA thatwas amplified in the same plate by the real-time RT-PCR. The resultsshown are the mean Ϯ SD of triplicate cultures, and the experiment wasrepeated 3 times with similar results (*P Ͻ .05; **P Ͻ .01).
IFN-␣ inhibited HCV RNA expression in Huh.8cells,20,21 we examined whether alcohol has a negative
Fig. 5. The role of NF-B in alcohol-induced HCV RNA expression in
Huh7 and Huh.8 cells. (A) Effect of alcohol on NF-B promoter in
impact on the anti-HCV effect of IFN-␣ in HCV repli-
pNF-B-Luc-transfected Huh7 and Huh.8 cells. Huh7 and Huh.8 cells
con containing cells. As expected, IFN-␣, when added to
(3 ϫ 105 cells/2 mL/well) plated in a 6-well plate were transfected with
Huh.8 cell cultures, significantly inhibited HCV RNA
pNF-B-Luc for 24 hours and then incubated with or without alcohol
expression (up to 90%) in Huh.8 cells (Fig. 4). This anti-
(100 mmol/L) and/or CAPE (20 g/mL) for 24 hours. NF-B promoter-directed luciferase activity quantitated from the cell-free lysates were
HCV effect of IFN-␣ on HCV, however, was attenuated
normalized using the total protein of the cell-free lysates. The data are
means Ϯ SD of triplicate cultures, and the experiment was repeated 3
Alcohol Activates NF-B. To investigate the possible
times with similar results (*P Ͻ .05; **P Ͻ .01). (B) Effect of CAPE onalcohol-enhanced HCV RNA expression in Huh.8 cells. Huh.8 cells (105
mechanism(s) responsible for alcohol-mediated upregula-
cells/mL/well) plated in a 24-well plate were incubated with or without
tion of HCV RNA expression, we examined whether al-
alcohol (100 mmol/L) and/or CAPE (20 g/mL) for 72 hours. Totalcellular RNA was extracted from the cell cultures and was then subjectedto real-time RT-PCR for HCV and GAPDH RNA quantification. The data areexpressed as HCV RNA copy numbers per reaction, which was normalizedbased on the ratio of HCV RNA/GAPDH mRNA that was amplified in thesame plate by the real-time RT-PCR. The results shown are the mean ϮSD of triplicate cultures, and the experiment was repeated 5 times withsimilar results (**P Ͻ .01).
cohol activates NF-B, an important transcriptionalnuclear factor that controls viral replication and cytokineproduction.28-30 Because HCV replicon expression inHuh.8 cells may activate NF-B, we transfected bothHuh7 cells, the parental cell line of Huh.8, and Huh.8cells with the plasmid (pNF-B-Luc) that contains
Fig. 4. Effect of alcohol on anti-HCV effect of IFN-␣ in Huh.8 cells.
NF-B promoter-linked to a luciferase gene. The trans-
Huh.8 cells (105 cells/mL/well) plated in a 24-well plate were incubated
fected cells (24 hours post-transfection) were then incu-
with or without alcohol (100 mmol/L) and/or IFN-␣ (100 U/mL) for 72hours. Total cellular RNA extracted from the cell cultures were subjected
bated with or without alcohol. Alcohol enhanced the
to real-time RT-PCR for HCV and GAPDH RNA quantification. The data are
NF-B promoter-directed luciferase activity in both
expressed as the percentage reduction in HCV RNA relative to the
Huh7 and Huh.8 cells (Fig. 5A). The basal levels of
untreated control levels (percent control). The results shown are the
mean Ϯ SD of triplicate cultures, and the experiment was repeated 5
B promoter-directed luciferase activity in Huh.8
times with similar results (**P Ͻ .01).
cells are higher than those in Huh7 cells (Fig. 5A). CAPE,
a specific inhibitor of the activation of NF-B, abrogatedalcohol-induced NF-B promoter activation (Fig. 5A) inboth Huh7 and Huh.8 cells. To further determinewhether NF-B activation is directly involved in alcohol-induced HCV RNA expression, we examined whetherCAPE abrogates the enhancing effect of alcohol in Huh.8cells. CAPE, when added to Huh.8 cell cultures, com-pletely blocked the enhancing effect of alcohol on HCVRNA expression (P Ͻ .01) (Fig. 5B). Naltrexone Abrogates Alcohol Action. Several lines
of evidence indicate that alcohol interferes with the en-dogenous opioid system.31-33 Morphine has the ability topromote HCV RNA expression in Huh.8 cells, which isblocked by naltrexone (Li Y, unpublished data). In addi-tion, we have documented that both Huh7 and Huh.8cells express -opioid receptor (Li Y, unpublished data). We therefore hypothesized that alcohol may activate theendogenous opioid system, which plays a role in alcohol-induced HCV RNA expression. To test this hypothesis,we first incubated Huh.8 cells with or without naltrexone(an opiate receptor antogonist) for 30 minutes before theaddition of alcohol to the Huh.8 cell cultures. Naltrexonenot only reversed the enhancing effect of alcohol on HCVRNA expression (Fig. 6A), but also inhibited the alcohol-
Fig. 6. Antagonizing effect of naltrexone on alcohol-induced NF-B
induced NF-B promoter activation (Fig. 6B).
promoter activation and HCV RNA expression in Huh.8 cells. (A) Effect of
4-MP and Cyanamide Inhibit Alcohol-Induced
naltrexone on alcohol-enhanced HCV RNA expression in Huh.8 cells. HCV RNA Expression. To determine whether the en-
Huh.8 cells (105 cells/mL/well) plated in a 24-well plate were incubatedwith naltrexone (10Ϫ8 mol/L) for 30 minutes before the addition of
hancing effect of alcohol on HCV RNA expression in
alcohol (100 mmol/L). Total cellular RNA extracted from the cell cultures
Huh.8 cells is specifically through the alcohol metabolite
was then subjected to real-time RT-PCR for HCV RNA quantification 72
acetaldehyde, we examined whether 4-MP, an inhibitor
hours after alcohol treatment. The data are expressed as HCV RNA copynumbers per reaction, which was normalized based on the ratio of HCV
of alcohol dehydrogenase, or cyanamide, an inhibitor of
RNA/GAPDH mRNA that was amplified in the same plate by the real-time
acetaldehyde dehydrogenase, blocks the enhancing effect
RT-PCR. The results shown are the mean Ϯ SD of triplicate cultures, and
of alcohol on HCV RNA expression in these cells. Pre-
the experiment was repeated 5 times with similar results (**P Ͻ .01).
treatment of Huh.8 cells with 4-MP or cyanamide atten-
(B) Effect of naltrexone on alcohol-induced NF-B promoter activation. Huh.8 cells (3 ϫ 105 cells/2 mL/well) plated in a 6-well plate were
uated the alcohol-induced increase of HCV RNA
transfected with pNF-B-Luc for 24 hours and then incubated with or
expression (Fig. 7), whereas the addition of 4-MP or cy-
without alcohol (100 mmol/L) and/or naltrexone (10Ϫ8 mol/L) for 24
anamide alone to the cell cultures had little impact on
hours. NF-B promoter-directed luciferase activity was normalized usingthe total protein of the cell-free lysates. The data are means Ϯ SD of
triplicate cultures, and the experiment was repeated 3 times with similarresults (**P Ͻ .01). Discussion
Alcohol abuse is a major cofactor in the development
of HCV associated liver disease.16-18 Chronic alcohol
number of responders in IFN therapy decreased as alcohol
abuse mediates liver damage as a result of increase in pro-
intake increased.40 These in vivo data strongly support the
duction of proinflammatory cytokines.34-37 In the setting
hypothesis that alcohol plays a role as a cofactor in pro-
of chronic HCV infection, alcohol ingestion has an addi-
moting HCV RNA expression. To directly link the action
tional effect of diminishing immune clearance and in-
of alcohol with HCV expression, we used recently devel-
creasing viral burden to hasten the onset of cirrhosis and
oped HCV replicon containing cell lines for this study.
hepatocellular carcinoma.38 Serum HCV RNA levels
The establishment of the subgenomic replicon system
were significantly higher in habitual alcohol drinkers with
represents major progress in the molecular biology of
chronic HCV infection than in infrequent alcohol drink-
HCV replication,21,27,41 which provides the first effective
ers with chronic HCV.39 HCV RNA levels were signifi-
cellular system available for the study of the dynamics of
cantly higher in alcohol drinkers than abstainers, and the
virus replication. Although the HCV replicon system
B. This finding is in agreement with a recent studyshowing that HCV NS5A alone or in the context of all theHCV nonstructural proteins is capable of inducing reac-tive oxygen species and activates NF-B.43 Interestingly,alcohol also has the ability to induce oxidative stress,which has been shown to activate NF-B.44 Thus, wehypothesized that the activation of NF-B is one of themechanisms by which alcohol enhances HCV replication. We showed that CAPE, a specific inhibitor of the activa-tion of NF-B,45 blocked alcohol-induced NF-B pro-moter activation (Fig. 5A), suggesting that the activationof NF-B may be involved in alcohol-mediated upregu-
Fig. 7. Effect of 4-MP, cyanamide on HCV RNA expression. Huh.8 cells
(105 cells/mL/well) plated in a 24-well plate were incubated with 4-MP
lation of HCV RNA expression. To confirm this specu-
(0.1 mol/L) or cyanamide (0.2 mol/L) for 30 minutes before the
lation, we examined whether CAPE could block the
addition of alcohol (100 mmol/L). Total cellular RNA extracted from the
enhancing effect of alcohol on HCV RNA expression.
cell cultures was then subjected to real-time RT-PCR for HCV and GAPDHRNA quantification 72 hours after alcohol treatment. The data are
Our data showed that CAPE, when added to Huh.8 cell
expressed as HCV RNA copy numbers per reaction, which was normalized
cultures, abolished the enhancing effect of alcohol on
based on the ratio of HCV RNA/GAPDH mRNA that was amplified in the
HCV RNA expression (Fig. 5B). These data indicate that
same plate by the real-time RT-PCR. The results shown are the mean Ϯ
SD of triplicate cultures, and the experiment was repeated 3 times with
similar results (**P Ͻ .01).
responsible for its action on HCV RNA expression.
Several lines of evidence indicate that alcohol interferes
with the endogenous opioid system,31-33 suggesting that
mimics only some aspects of HCV replication, this cell
the endogenous opioid system may be involved in alcohol
system provides an important means of investigating viral
action. The endogenous opioids play a key role in the
RNA and protein synthesis21,27,41 and of characterizing
rewarding (addictive) properties of alcohol.46,47 Pretreat-
those factor(s) that regulate HCV RNA expression. Using
ment with opiate receptor antagonists (e.g., Naltrexone)
this system, we have for the first time shown that alcohol
decreases alcohol consumption in humans.48,49 Further-
enhanced HCV replicon expression at both mRNA and
more, treatment with naltrexone, an opiate antagonist,
protein levels (Figs. 1 through 3). This enhancing effect of
decreases ethanol consumption by alcoholic subjects.47-50
alcohol is mediated specifically through its metabolite ac-
Opioid content, biosynthesis, and release are altered after
etaldehyde, because 4-MP or cyanamide, the specific in-
alcohol treatment, although there is considerable dis-
hibitors of alcohol metabolism, blocks alcohol action on
agreement regarding the direction of changes in the activ-
HCV RNA expression (Fig. 7). These data indicate that
ity of the endogenous opioid system by alcohol.46 We
Huh.8 cells are able to metabolize alcohol, probably
recently showed that Huh.8 cells express -opioid recep-
through alcohol dehydrogenase, the main enzyme respon-
tor and that morphine enhances HCV RNA expression in
these cells (Li Y, unpublished data). Thus, we hypothesize
The underlying mechanisms of interaction between
that alcohol may activate the endogenous opioid system,
alcohol and HCV remain largely unknown. In the present
through which it induces HCV RNA expression. We
study, we also investigated the possible mechanism(s) re-
showed that naltrexone, an opiate receptor antagonist,
sponsible for alcohol-mediated upregulation of HCV
not only blocked the promoting effect of alcohol on HCV
RNA expression. Because NF-B is a critical nuclear tran-
RNA expression in Huh.8 cells (Fig. 6A), but also dimin-
scription factor involved in the activation of viral replica-
ished the enhancing effect of alcohol on NF-B promoter
tion and cytokine production28-30 and alcohol enhances
activation in these cells (Fig. 6B). These data strongly
NF-B activation in different cell systems, such as
suggest that the activation of the endogenous opioid sys-
HepG2 cells,42 we investigated whether alcohol, through
tem is also implicated in alcohol-induced HCV RNA ex-
the activation of NF-B, enhances HCV expression in
Huh7 and Huh.8 cells. We showed that alcohol enhanced
Clinical trials indicate a therapeutic benefit of IFN-␣
NF-B promoter-directed luciferase activity in both
treatment in chronic HCV infection.13,18 Currently avail-
Huh7 and Huh.8 cells (Fig. 5A). Because the basal levels
able combination therapy with IFN-␣ and ribavirin is,
of NF-B promoter-directed luciferase activity in Huh.8
however, effective in less than 50% of treated sub-
cells are higher than those in Huh7 cells, it seems that
jects.7,10,11 Although a history of alcohol abuse is not a
HCV replicon expression in Huh.8 cells can activate NF-
contraindication to clinical therapy, continued alcohol
use during therapy adversely affects response to HCV
References
treatment. Heavy alcohol consumption reduces the effi-
1. Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M.
cacy of IFN-␣ therapy for chronic HCV infection,14 and
Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral
this adverse effect of alcohol drinking on efficacy might be
hepatitis genome. Science 1989;244:359-362.
reversed, partly, by abstinence for a long period before
2. Kuo G, Choo QL, Alter HJ, Gitnick GL, Redeker AG, Purcell RH, Miya-
mura T, et al. An assay for circulating antibodies to a major etiologic virus
treatments. Thus, it is important to identify whether al-
of human non-A, non-B hepatitis. Science 1989;244:362-364.
cohol is one of the cofactors that are responsible for the
3. Strader DB, Seeff LB. The natural history of chronic hepatitis C infection.
failure of IFN-␣ treatment. The HCV replicon system
Eur J Gastroenterol Hepatol 1996;8:324-328.
4. Alter MJ, Kruszon-Moran D, Nainan OV, McQuillan GM, Gao F, Moyer
has been successfully used to examine the anti-HCV effect
LA, Kaslow RA, et al. The prevalence of hepatitis C virus infection in the
of IFN-␣.20,21 IFN-␣ inhibits HCV RNA expression in
United States, 1988 through 1994. N Engl J Med 1999;341:556-562.
Huh.8 cells, as shown by a declined HCV RNA expres-
5. Alter MJ. Epidemiology of hepatitis C in the West. Semin Liver Dis 1995;
sion over time.21 Our data (Fig. 4) showing that IFN-␣
6. Choo QL, Richman KH, Han JH, Berger K, Lee C, Dong C, Gallegos C,
significantly inhibited (up to 90%) HCV RNA expres-
et al. Genetic organization and diversity of the hepatitis C virus. Proc Natl
sion in Huh.8 cells further confirms this observation.
7. Alter HJ, Seeff LB. Recovery, persistence, and sequelae in hepatitis C virus
Thus, the Huh.8 cell line is an excellent in vitro model for
infection: a perspective on long-term outcome. Semin Liver Dis 2000;20:
studying whether alcohol interferes with the anti-HCV
effect of IFN-␣ on HCV RNA expression.21 We hypoth-
8. Alter MJ, Margolis HS, Krawczynski K, Judson FN, Mares A, Alexander
esized that alcohol abuse may have a negative impact on
WJ, Hu PY, et al. The natural history of community-acquired hepatitis Cin the United States. The Sentinel Counties Chronic non-A, non-B Hep-
the anti-HCV effect of IFN-␣. Our data support this
atitis Study Team. N Engl J Med 1992;327:1899-1905.
hypothesis, showing that alcohol compromised the anti-
9. Kiyosawa K, Sodeyama T, Tanaka E, Gibo Y, Yoshizawa K, Nakano Y,
HCV effect of IFN-␣ in Huh.8 cells (Fig. 4). This finding
Furuta S, et al. Interrelationship of blood transfusion, non-A, non-B hep-atitis and hepatocellular carcinoma: analysis by detection of antibody to
suggests the possibility that alcohol may reduce efficacy of
hepatitis C virus. HEPATOLOGY 1990;12:671-675.
IFN-␣ therapy in vivo. The mechanism(s) responsible for
10. Davis GL, Esteban-Mur R, Rustgi V, Hoefs J, Gordon SC, Trepo C,
the IFN-␣–mediated therapeutic effect remain unclear.20
Shiffman ML, et al. Interferon alfa-2b alone or in combination with riba-virin for the treatment of relapse of chronic hepatitis C. International
Thus, further studies are critical to determine the mech-
Hepatitis Interventional Therapy Group. N Engl J Med 1998;339:1493-
anism(s) responsible for the anti-HCV ability of IFN-␣
and whether alcohol has the ability to interfere with the
11. McHutchison JG, Gordon SC, Schiff ER, Shiffman ML, Lee WM, Rustgi
VK, Goodman ZD, et al. Interferon alfa-2b alone or in combination with
mechanism(s) involved in IFN-␣ action against HCV.
ribavirin as initial treatment for chronic hepatitis C. Hepatitis Interven-
In summary, our data show that alcohol has the ability
tional Therapy Group. N Engl J Med 1998;339:1485-1492.
to potentiate HCV replicon expression at both mRNA
12. Cook RT. Alcohol abuse, alcoholism, and damage to the immune sys-
tem—a review. Alcohol Clin Exp Res 1998;22:1927-1942.
and protein levels. Also, alcohol compromises the anti-
13. Corrao G, Arico S. Independent and combined action of hepatitis C virus
HCV effect of IFN-␣ in HCV replicon containing he-
infection and alcohol consumption on the risk of symptomatic liver cir-
patic cells. NF-B activation by alcohol may be, at least
partially, responsible for alcohol-mediated upregulation
14. Ohnishi K, Matsuo S, Matsutani K, Itahashi M, Kakihara K, Suzuki K, Ito
S, et al. Interferon therapy for chronic hepatitis C in habitual drinkers:
of HCV RNA expression. In addition, alcohol-induced
comparison with chronic hepatitis C in infrequent drinkers. Am J Gastro-
activation of the endogenous opioid system may be an
additional mechanism by which alcohol induces HCV
15. Okazaki T, Yoshihara H, Suzuki K, Yamada Y, Tsujimura T, Kawano K,
Abe H. Efficacy of interferon therapy in patients with chronic hepatitis C.
expression. These important observations in conjunction
Comparison between non-drinkers and drinkers. Scand J Gastroenterol
with the in vivo observations13-18,40 are beneficial for fur-
ther biological and immunological studies on cellular and
16. Pessione F, Degos F, Marcellin P, Duchatelle V, Njapoum C, Martinot-
Peignoux M, Degott C, et al. Effect of alcohol consumption on serum
molecular mechanism(s) involved in regulation of HCV
hepatitis C virus RNA and histological lesions in chronic hepatitis C.
replication in both in vitro and in vivo models. Our data
also provide practical guidance toward the reduction of
17. Regev A, Jeffers LJ. Hepatitis C and alcohol. Alcohol Clin Exp Res 1999;
risk factors that interfere with IFN-␣– based therapy
18. Wiley TE, McCarthy M, Breidi L, Layden TJ. Impact of alcohol on the
histological and clinical progression of hepatitis C infection. HEPATOLOGY1998;28:805-809.
19. Szabo G. Consequences of alcohol consumption on host defence. Alcohol
Rice (The Rockefeller University) for generously provid-
ing Huh8 and Huh7 cell lines. The authors also are grate-
20. Guo JT, Bichko VV, Seeger C. Effect of alpha interferon on the hepatitis C
virus replicon. J Virol 2001;75:8516-8523.
ful to Dr. Christoph Seeger (Fox Chase Cancer Center)
21. Blight KJ, Kolykhalov AA, Rice CM. Efficient initiation of HCV RNA
replication in cell culture. Science 2000;290:1972-1974.
22. Yang JH, Lai JP, Douglas SD, Metzger D, Zhu XH, Ho WZ. Real-time
37. Deviere J, Content J, Denys C, Vandenbussche P, Schandene L, Wybran J,
RT-PCR for quantitation of hepatitis C virus RNA. J Virol Methods
Dupont E. High interleukin-6 serum levels and increased production by
leucocytes in alcoholic liver cirrhosis. Correlation with IgA serum levels
23. Szabo G, Catalano D, Bellerose G, Mandrekar P. Interferon alpha and
and lymphokines production. Clin Exp Immunol 1989;77:221-225.
alcohol augment nuclear regulatory factor-kappaB activation in HepG2
38. Prakash O, Mason A, Luftig RB, Bautista AP. Hepatitis C virus (HCV)
cells, and interferon alpha increases pro-inflammatory cytokine produc-
and human immunodeficiency virus type 1 (HIV-1) infections in alcohol-
tion. Alcohol Clin Exp Res 2001;25:1188-1197.
24. Petrak D, Memon SA, Birrer MJ, Ashwell JD, Zacharchuk CM. Domi-
39. Oshita M, Hayashi N, Kasahara A, Hagiwara H, Mita E, Naito M, Kata-
nant negative mutant of c-Jun inhibits NF-AT transcriptional activity and
yama K, et al. Increased serum hepatitis C virus RNA levels among alco-
prevents IL-2 gene transcription. J Immunol 1994;153:2046-2051.
holic patients with chronic hepatitis C. HEPATOLOGY 1994;20:1115-1120.
25. Pierce JW, Lenardo M, Baltimore D. Oligonucleotide that binds nuclear
40. Loguercio C, Di Pierro M, Di Marino MP, Federico A, Disalvo D, Crafa
factor NF-kappa B acts as a lymphoid-specific and inducible enhancer
E, Tuccillo C, et al. Drinking habits of subjects with hepatitis C virus–
element. Proc Natl Acad Sci U S A 1988;85:1482-1486.
related chronic liver disease: prevalence and effect on clinical, virological
26. Luckow B, Schutz G. CAT constructions with multiple unique restriction
and pathological aspects. Alcohol Alcohol 2000;35:296-301.
sites for the functional analysis of eukaryotic promoters and regulatory
41. Rosenberg S. Recent advances in the molecular biology of hepatitis C virus.
elements. Nucleic Acids Res 1987;15:5490.
27. Lohmann V, Korner F, Koch J, Herian U, Theilmann L, Bartenschlager R.
42. Kim WH, Hong F, Jaruga B, Hu Z, Fan S, Liang TJ, Gao B. Additive
Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line.
activation of hepatic NF-kappaB by ethanol and hepatitis B protein X(HBX) or HCV core protein: involvement of TNF-alpha receptor 1-inde-
pendent and -dependent mechanisms. FASEB J 2001;15:2551-2553.
28. Powell JD, Bednarik DP, Folks TM, Jehuda-Cohen T, Villinger F, Sell
43. Gong G, Waris G, Tanveer R, Siddiqui A. Human hepatitis C virus NS5A
KW, Ansari AA. Inhibition of cellular activation of retroviral replication by
protein alters intracellular calcium levels, induces oxidative stress, and ac-
CD8ϩ T cells derived from non-human primates. Clin Exp Immunol
tivates STAT-3 and NF-kappa B. Proc Natl Acad Sci U S A 2001;98:9599-
29. Swingler S, Morris A, Easton A. Tumour necrosis factor alpha and inter-
44. Bailey SM, Cunningham CC. Acute and chronic ethanol increases reactive
leukin-1 beta induce specific subunits of NFKB to bind the HIV-1 en-
oxygen species generation and decreases viability in fresh, isolated rat hepa-
hancer: characterisation of transcription factors controlling human
tocytes. HEPATOLOGY 1998;28:1318-1326.
immunodeficiency virus type 1 gene expression in neural cells. Biochem
45. Natarajan K, Singh S, Burke TR, Jr., Grunberger D, Aggarwal BB. Caffeic
Biophys Res Commun 1994;203:623-630.
acid phenethyl ester is a potent and specific inhibitor of activation of
30. Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell 2002;
nuclear transcription factor NF-kappa B. Proc Natl Acad Sci U S A 1996;
31. O’Brien CP, Volpicelli LA, Volpicelli JR. Naltrexone in the treatment of
46. Herz A. Endogenous opioid systems and alcohol addiction. Psychophar-
alcoholism: a clinical review. Alcohol 1996;13:35-39.
32. Reid LD. Endogenous opioids and alcohol dependence: opioid alkaloids
47. Gianoulakis C. Implications of endogenous opioids and dopamine in al-
and the propensity to drink alcoholic beverages. Alcohol 1996;13:5-11.
coholism: human and basic science studies. Alcohol Alcohol 1996;31:33-
33. Van Ree JM. Endorphins and experimental addiction. Alcohol 1996;13:
48. Volpicelli JR, Alterman AI, Hayashida M, O’Brien CP. Naltrexone in the
34. Lin HZ, Yang SQ, Zeldin G, Diehl AM. Chronic ethanol consumption
treatment of alcohol dependence.[comment]. Arch Gen Psychiatry 1992;
induces the production of tumor necrosis factor-alpha and related cyto-
kines in liver and adipose tissue. Alcohol Clin Exp Res 1998;22:231S-
49. O’Malley SS, Jaffe AJ, Chang G, Schottenfeld RS, Meyer RE, Rounsaville
B. Naltrexone and coping skills therapy for alcohol dependence. A con-
35. McClain CJ, Cohen DA. Increased tumor necrosis factor production by
trolled study. Arch Gen Psychiatry 1992;49:881-887.
monocytes in alcoholic hepatitis. HEPATOLOGY 1989;9:349-351.
50. Jaffe AJ, Rounsaville B, Chang G, Schottenfeld RS, Meyer RE, O’Malley
36. Khoruts A, Stahnke L, McClain CJ, Logan G, Allen JI. Circulating tumor
SS. Naltrexone, relapse prevention, and supportive therapy with alcohol-
necrosis factor, interleukin-1 and interleukin-6 concentrations in chronic
ics: an analysis of patient treatment matching. J Consult Clin Psychol
alcoholic patients. HEPATOLOGY 1991;13:267-276.
Get Plastic Out Of Your Diet our foods—plastics, pesticides, growth hormones, irradiation, and microwave. This is clear from the mass of expert and citizen testimony against such technologies that In the interests of a healthy population, we regulatory agencies bend over backwards and have decided to re-print this well jump through flaming hoops to please their researched and irr
j o u r n a l h o m e p a g e : w w w. e l s ev i e r. c o m / l o c a t e / s c i to t e n vAssessment of DNA damage in coal open-cast mining workers using thecytokinesis-blocked micronucleus test and the comet assayGrethel León-Mejía Lyda Espitia-Pérez , Luz Stella Hoyos-Giraldo , Juliana Da Silva ,Andreas Hartmann , João Antônio Pêgas Henriques ,, Milton Quintana a Laboratorio de In
 Effect of Alcohol on HCV Replicon. To evaluate the
Effect of Alcohol on HCV Replicon. To evaluate the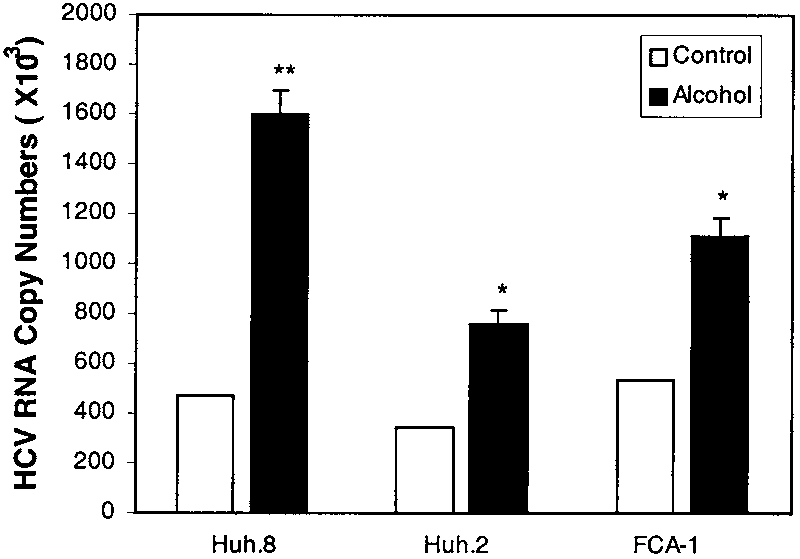
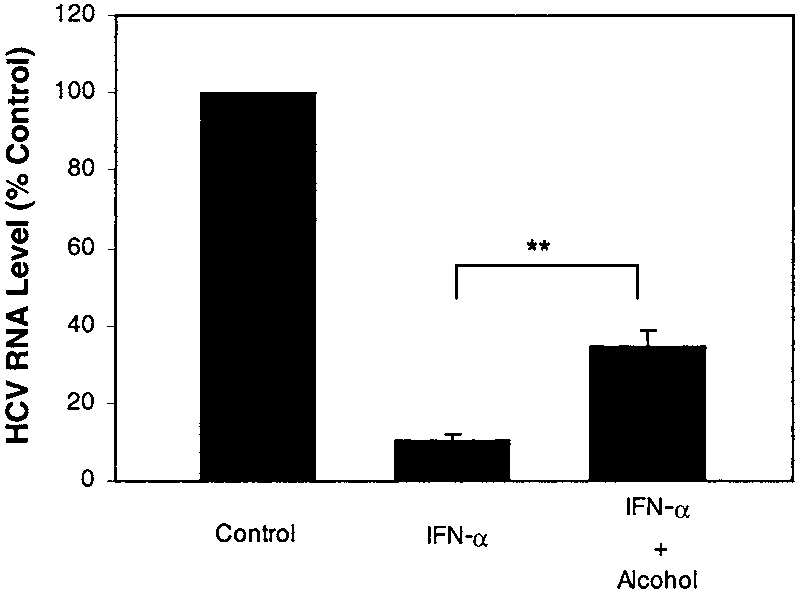
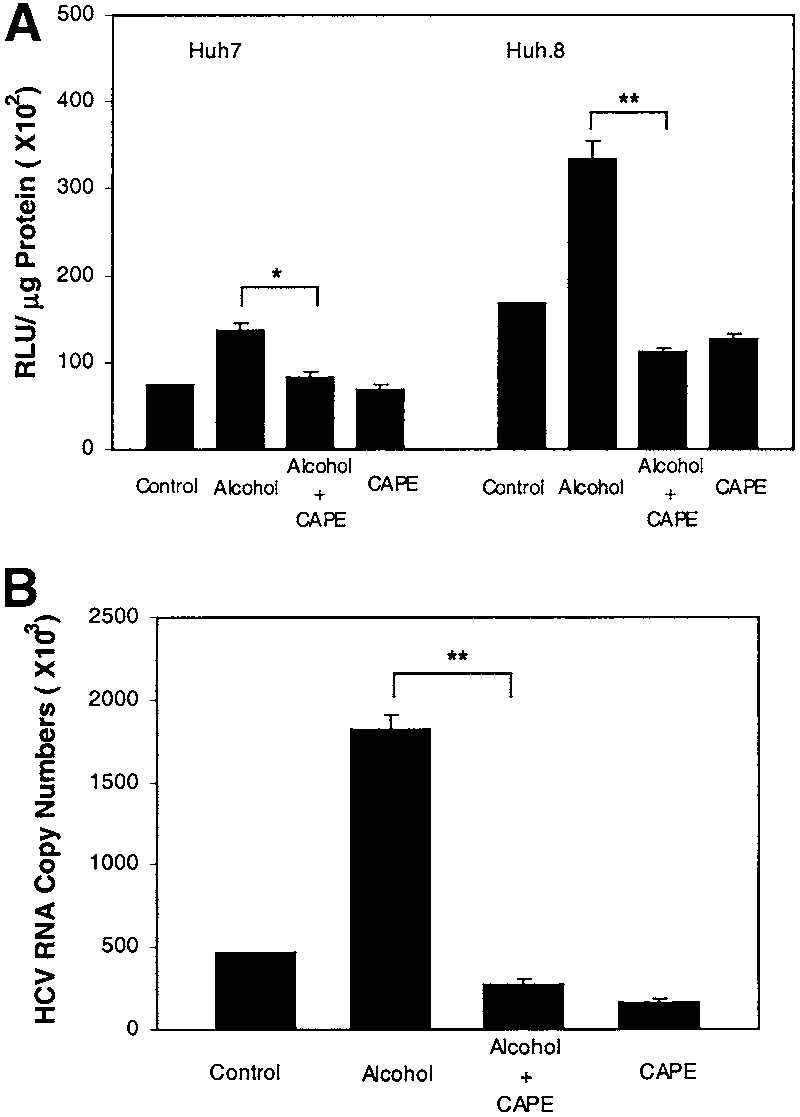 Fig. 3. Effect of alcohol on HCV RNA expression in different HCV
replicon cell clones. Huh.8, Huh.2, and FCA-1 cells (105 cells/mL/well)plated in a 24-well plate were incubated with or without alcohol (100mmol/L) for 72 hours. Total cellular RNA extracted from the cell cultureswas subjected to real-time RT-PCR for HCV and GAPDH RNA quantifica-tion. The data are expressed as HCV RNA copy numbers per reaction,which was normalized based on the ratio of HCV RNA/GAPDH mRNA thatwas amplified in the same plate by the real-time RT-PCR. The resultsshown are the mean Ϯ SD of triplicate cultures, and the experiment wasrepeated 3 times with similar results (*P Ͻ .05; **P Ͻ .01).
Fig. 3. Effect of alcohol on HCV RNA expression in different HCV
replicon cell clones. Huh.8, Huh.2, and FCA-1 cells (105 cells/mL/well)plated in a 24-well plate were incubated with or without alcohol (100mmol/L) for 72 hours. Total cellular RNA extracted from the cell cultureswas subjected to real-time RT-PCR for HCV and GAPDH RNA quantifica-tion. The data are expressed as HCV RNA copy numbers per reaction,which was normalized based on the ratio of HCV RNA/GAPDH mRNA thatwas amplified in the same plate by the real-time RT-PCR. The resultsshown are the mean Ϯ SD of triplicate cultures, and the experiment wasrepeated 3 times with similar results (*P Ͻ .05; **P Ͻ .01).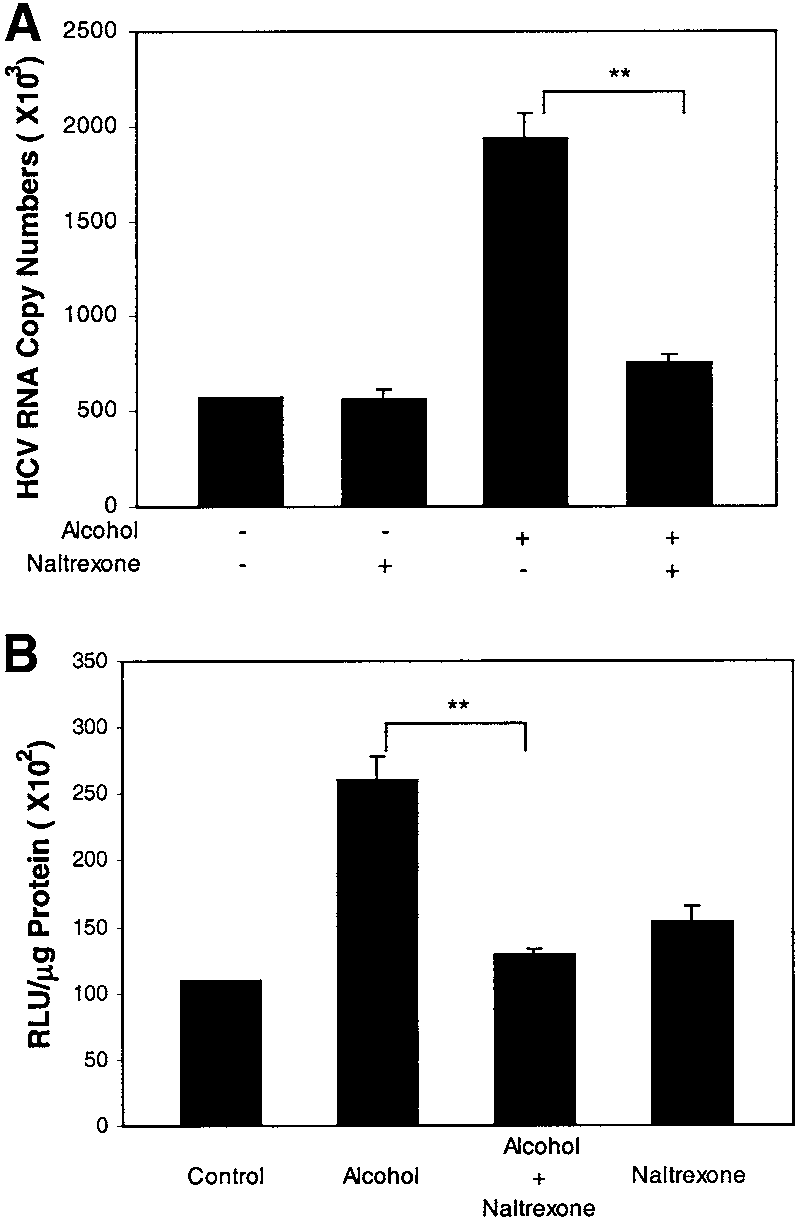 a specific inhibitor of the activation of NF-B, abrogatedalcohol-induced NF-B promoter activation (Fig. 5A) inboth Huh7 and Huh.8 cells. To further determinewhether NF-B activation is directly involved in alcohol-induced HCV RNA expression, we examined whetherCAPE abrogates the enhancing effect of alcohol in Huh.8cells. CAPE, when added to Huh.8 cell cultures, com-pletely blocked the enhancing effect of alcohol on HCVRNA expression (P Ͻ .01) (Fig. 5B).
a specific inhibitor of the activation of NF-B, abrogatedalcohol-induced NF-B promoter activation (Fig. 5A) inboth Huh7 and Huh.8 cells. To further determinewhether NF-B activation is directly involved in alcohol-induced HCV RNA expression, we examined whetherCAPE abrogates the enhancing effect of alcohol in Huh.8cells. CAPE, when added to Huh.8 cell cultures, com-pletely blocked the enhancing effect of alcohol on HCVRNA expression (P Ͻ .01) (Fig. 5B).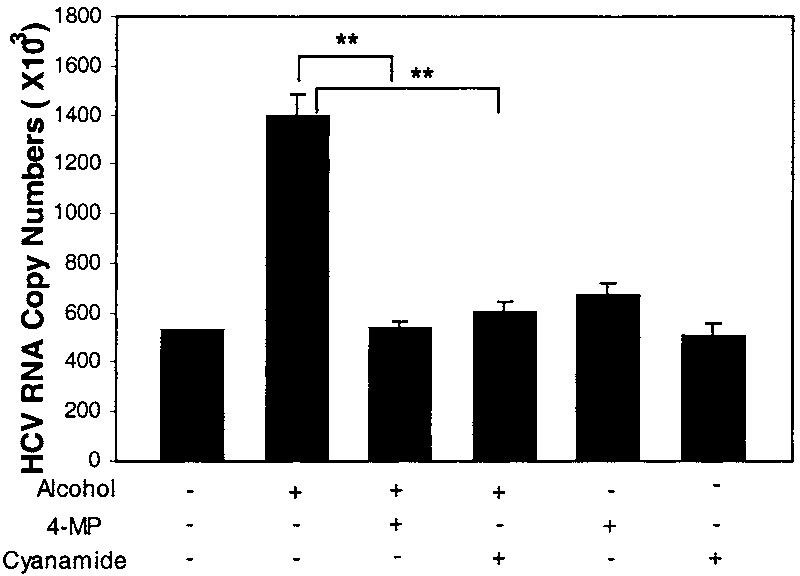 B. This finding is in agreement with a recent studyshowing that HCV NS5A alone or in the context of all theHCV nonstructural proteins is capable of inducing reac-tive oxygen species and activates NF-B.43 Interestingly,alcohol also has the ability to induce oxidative stress,which has been shown to activate NF-B.44 Thus, wehypothesized that the activation of NF-B is one of themechanisms by which alcohol enhances HCV replication.
B. This finding is in agreement with a recent studyshowing that HCV NS5A alone or in the context of all theHCV nonstructural proteins is capable of inducing reac-tive oxygen species and activates NF-B.43 Interestingly,alcohol also has the ability to induce oxidative stress,which has been shown to activate NF-B.44 Thus, wehypothesized that the activation of NF-B is one of themechanisms by which alcohol enhances HCV replication.