Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
In vitro effects of 2-methoxyestradiol on cell numbers, morphology, cell cycle progression, and apoptosis induction in oesophageal carcinoma cells
Cell Biochem Funct 2009; 27: 205–210. Published online 2 April 2009 in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/cbf.1557
In vitro effects of 2-methoxyestradiol on cell numbers, morphology,cell cycle progression, and apoptosis induction in oesophagealcarcinoma cells
Veneesha Thaver 1,2, Mona-Liza Lottering 2, Dirk van Papendorp 2 and Annie Joubert 2*
1Department of Physiology, University of Limpopo, Garankuwa, Pretoria, South Africa2Department of Physiology, University of Pretoria, Pretoria, South Africa
The influence of 2-methoxyestradiol (2-ME) was investigated on cell numbers, morphology, cell cycle progression, and apoptosis induction inan oesophageal carcinoma cell line (WHCO3). Dose-dependent studies (1 Â 10À9M–1 Â 10À6M) revealed that 2-ME significantly reducedcell numbers to 60% in WHCO3 after 72 h of exposure at a concentration of 1 Â 10À6M compared to vehicle-treated cells. Morphologicalstudies entailing light-, fluorescent-, as well as transmission electron microscopy (TEM) confirmed 2-ME’s antimitotic effects. These resultsindicated hallmarks of apoptosis including cell shrinkage, hypercondensation of chromatin, cell membrane blebbing, and apoptotic bodies intreated cells. Flow cytometric analyses demonstrated an increase in the G2/M-phase after 2-ME exposure; thus preventing cells fromproceeding through the cell cycle. b-tubulin immunofluorescence revealed that 2-ME caused spindle disruption. In addition, increasedexpression of death receptor 5 protein was observed further supporting the proposed mechanism of apoptosis induction via the extrinsicpathway in 2-ME-exposed oesophageal carcinoma cells. Copyright # 2009 John Wiley & Sons, Ltd.
key words — oesophageal carcinoma; 2-methoxyestradiol; metaphase block; apoptosis
phosphorylation and inactivation of Bcl-2, a pro-apoptoticprotein, thus contributing to apoptotic induction.8,9 Never-
2-Methoxyestradiol (2-ME), a 17-beta estradiol metabolite,
theless, apoptotic induction via the extrinsic and intrinsic
is a mitogen antagonist and tubulin poison that hinders cell
pathways appears to be dependent on cell type.13–15
proliferation and induces apoptosis in a large diversity of
It is also known that 2-ME displays a dose-dependent
non-tumor and tumor cells.1–4 2-ME implements both its
biphasic pattern on cell proliferation at concentrations
antiangiogenic and antitumor influence regardless of the
ranging from 10À8 to 10À5 M. Stimulatory effects have been
cell’s hormone receptor status and is accountable for
demonstrated at low concentrations of 2-ME and inhibitory
abnormal mitotic spindle formation and mitotic accumu-
effects were observed at high concentrations.3,11,16 Corre-
lation in both estrogen receptor (ER) positive- and ER-
spondingly, in vivo studies have demonstrated stimulation
negative cells.5–7 Accordingly, this endogenous estradiol
and inhibition of tumor growth by 2-ME depending on
metabolite has manifested as a potential anticancer agent.5
dosage.17,18 In concert, these research studies imply a
Current evidence has suggested that 2-ME is the causative
multifaceted nature of the action of 2-ME. Data illustrated
agent leading to an increase in Cdc2 kinase activity, the
that, in addition to the established signaling pathways there
activation of c-Jun NH2-terminal kinase signaling, gener-
may be supplementary pathways that have not been
ation of reactive oxygen species and an altered ratio of Bax/
Bcl-2 in favor of Bax, ultimately culminating into
ME.13,14,16,19–21 This biphasic effect may also be cell line
apoptosis.8–11 Cell division cycle (Cdc) 2 kinase activity
is a cell cycle regulatory component essential for
Preclinical data illustrated that 2-ME might be considered
commencement of mitosis, whereas Cdc2 inactivation is
in the treatment of multiple myeloma, sarcoma and other
needed for mitotic exit. Prolonged Cdc2 activity can sustain
solid tumors, therefore portraying it as a possible anticancer
the cell in mitosis for an indefinite period until particular
agent when compared to conventional chemotherapeutic
conditions are met for mitotic exit.12 JNK is involved in the
treatments.5,16,22,23 Phase I and phase II clinical trials with 2-ME revealed its therapeutic potential when administered topatients with metastatic breast cancers and prostate cancers
* Correspondence to: A. Joubert, Department of Physiology, P.O. Box 2034,
with only minor side-effects in some of the patients namely
Pretoria 0001, South Africa. Tel: þ27 12 3192246; Fax: þ27 12 3211679. E-mail: annie.joubert@up.ac.za
hot flushes, reversible liver enzyme elevations, fatigue, and
Copyright # 2009 John Wiley & Sons, Ltd.
diarrhoea.5,16,19,22 Clinical studies employing 2-ME in
(v/v). Controls included showed that 0.1% had no toxic
cancer patients revealed that treatment is linked with
effects on these cells in experiments conducted.
clinical advantages including prolonged, stable disease,partial or complete responses, and an exceptional safety
Since the mechanism of action of 2-ME is multifaceted
Exponentially growing WHCO3 cells were seeded in 24-
and appears to vary according to cell type2,5,11,24 the aim of
well culture plates at a density of 20 000 viable cells per well
this study was to investigate the mechanism of action of 2-
and exposed to a dilution series of 2-ME with a final
ME in an oesophageal carcinoma cell line by determining its
concentration of 10À6, 10À7, 10À8, 10À9 M respectively for
influence on cell numbers, morphology, cell cycle pro-
72 h at 378C. The experiment was terminated by replacing
the growth medium with 300ml of 1% glutaraldehyde in PBSfor 15 min. Crystal violet (1%, in PBS) was added for30 min. The culture wells were subsequently immersed in
running tap water for 15 min. After the plates had dried,
500 ml of 0.2% Triton X-100 was added to each well. Plateswere incubated for 90 min and 200 ml of the liquid content
2-ME, Eagles’ Minimum Essential Medium with Earle’s
was transferred to 96-well plates.25 The absorbance
salts, L-glutamine and NaHCO3 (MEM), Trypsin-EDTA,
(measured at 570 nm) of the samples was analyzed using
trypan blue, thymidine, hydroxyurea, anti-human Bcl-2
antibody, mouse monoclonal antibody against human b-
X800 Universal Microplate Reader (Bio-Tek Instru-
ments Inc., Analytical Diagnostic Products, Weltevreden
tubulin (Clone 2-28-33) biotin-conjugated anti-mouse IgG
SA). Results shown are representative of three independent
(Fab-specific, developed in goat), FITC-conjugate diluent
experiments (each conducted in triplicate).
and ExtrAvidin1-FITC conjugate were supplied by SigmaChemical Co. (St. Louis, MO, USA). Hematoxylin, eosin,
ethanol, xylol, and Entellan fixative were purchased from
Merck (Darmstadt, Germany). Propidium iodide was
Propidium iodide and hoechst 33342 staining. To study the
supplied by DAKO Chemical Supplies (Glostrup, Den-
viability and presence of apoptotic cells after 2-ME
mark). DAKO LSAB Kit was purchased from Dako
treatment, 500 000 WHCO3 cells were seeded onto heat-
Corporation (Santa Barbara, CA, USA). The death receptor
sterilized cover slips and exposed to 1 Â 10À6 M 2-ME for
five antibody and human anti-goat IgG were purchased from
24 h. An exposure time of 24 h was chosen since significant
Calbiochem (Darmstadt, Germany). Heat-inactivated fetal
reductions in cell number were visible after 24 h of treatment
calf serum (FCS), sterile cell culture flasks and plates were
with 2-ME. Medium was removed, the cells were gently
obtained though Sterilab Services (Kempton Park, Johan-
rinsed with PBS and 2 ml of 0.5 mg mlÀ1 Hoechst 33342
nesburg, SA). Phosphate buffered saline (PBS), penicillin,
(HO) in PBS was added to each well. Samples were
streptomycin, and fungizone were obtained from Highveld
Biological (Sandringham, SA). Quetol, Reynolds’ lead
0.5 ml of propidium iodide (PI) solution (40 mg mlÀ1 in
citrate, aqueous uranyl acetate, and toludine blue were
PBS) was added. Within 5 min cover slips were mounted on
purchased from Merck Co. (Johannesburg, South Africa).
microscope slides with mounting fluid (90% glycerol, 4% N-
All other chemicals were of analytical grade and supplied by
propyl-gallate, 6% PBS) and examined under a fluorescence
Sigma Chemical Co. (St. Louis, MO, USA).
microscope. Photographs were taken with 400 ASA film ona Nikon Optiphot microscope (Nikon, Tokyo, Japan) withUV-light and a blue filter. Viable and apoptotic (although
having irregular appearances) cells will stain light blue. The
WHCO3 cells were a gift from Professors Veale and
latter phenomenon illustrates that these cells have functional
Thornley (Department of Zoology, University of Witwa-
cell membranes capable of excluding PI. However, cells
tersrand, Johannesburg, South Africa). These cells were
with compromised membrane integrity will stain bright
obtained though a biopsy from a patient with squamous
oesophageal carcinoma and are described as a poorlydifferentiated, non-keratinizing
Hematoxylin and eosin staining (H and E staining)
propagated as monolayers in MEM at 378C in a humidifiedatmosphere containing 5% CO2. Media were supplemented
WHCO3 cells (250 000) were seeded onto heat-sterilized
with 10% heat-inactivated fetal calf serum, penicillin
cover slips in 6-well plates. Cells were exposed to
(100 mg LÀ1), streptomycin (100 mg LÀ1), and fungizone
1 Â 10À6 M 2-ME for a period of 24 h at 378C. Many cells
(250 mg LÀ1). Non-viable cells were excluded with the
were not adherent to cover slips after exposures of 24–48 h
trypan blue staining procedure. Stock solutions of 2-ME
and had disintegrated to become floating debris. Thus, it was
were prepared in dimethyl sulphoxide (DMSO) at concen-
not possible to observe morphological changes occurring
trations of 2 Â 10À3M and stored at room temperature. The
during this period when studying the influence of 2-ME on
DMSO content of the final dilutions never exceeded 0.1%
WHCO3 cells by means of H and E staining. Cells were
Copyright # 2009 John Wiley & Sons, Ltd.
Cell Biochem Funct 2009; 27: 205–210.
INFLUENCE OF 2-ME IN OESOPHAGEAL CARCINOMA CELLS
fixed in Bouin’s fixative for 60 min after exposure to 2-ME
1 h (1:100). Following washing, cells were finally incubated
and stained by standard hematoxylin and eosin staining
with ExtrAvidin1-FITC conjugate (1:200 in FITC-con-
jugate diluent) for 1 h. The cover slips were mounted with aglycerol-based mounting fluid after the final 3 Â 5-min washstep. The cells were examined with a Nikon Optiphot
microscope equipped with an episcopic-fluorescence attach-
Exponentially growing cells (500 000) were seeded in
ment and an excitation-emission filter with an average
25 cm2 flasks and exposed to 0.1% DMSO (vehicle control)
and 1 Â 10À6 M 2-ME for 24 h respectively. Cells werewashed with PBS (3x) and scraped off the bottom of theflask. Subsequently, ultra-thin sections of cells were
Immunofluorescent detection of death receptor 5 (DR5)
prepared. Cells were fixed in 2.5% glutaraldehyde in
Cells (500 000) were seeded onto heat-sterilized glass cover
0.075 M phosphate buffer, (pH 7.4–7.6) for 1 h and rinsed
slips in 6-well plates and exposed to 2-ME or DMSO
3 times for 5 min each with 0.075 M phosphate buffer.
controls for a period of 24 h at 378C. The cells were then
Thereafter sections were fixed in 0.25% aqueous osmium
fixed in 10% formalin (2 mM EGTA in PBS) for 10 min and
tetroxide and rinsed (3x) in distilled water in a fume hood.
permeabilized in ice-cold 97% methanol containing 2 mM
Samples were dehydrated in ethanol (70, 100%), infiltrated
EGTA at -208C for 10 min. Cells were subsequently washed
with 30% quetol in acetone for 1 h and furthermore
in PBS (3 Â 5 min) before incubation for 1 h with a mouse
infiltrated with 60% quetol in ethanol for 1 h, and thereafter
monoclonal antibody against human Death Receptor
with pure quetol for 4 h. Sections were polymerized at 658C
5 (Clone 2-28-33; 1:1000). After washing with PBS, the
for 24–36 h. Ultra-thin sections were mounted on grids,
cells were incubated with biotin-conjugated anti-mouse IgG
contrasted for 10 min in 4% aqueous uranyl acetate and
(Fab-specific, developed in goat) in FITC-conjugate diluent
rinsed in water. Enhancement of contrast was obtained by
as secondary antibody for 1 h (1:100). Following washing,
placing the samples in Reynolds’ lead citrate for 2 min and
cells were finally incubated with ExtrAvidin1-FITC
rinsing the samples in water. Samples were cut into 0.5 mm
conjugate (1:200 in FITC-conjugate diluent) for 1 h. The
monitor sections, stained with toluidine blue, and immersed
cover slips were mounted with a glycerol-based mounting
fluid after the final 3 Â 5-min wash step. Cells wereexamined with a Nikon Optiphot microscope equipped
with an episcopic-fluorescence attachment and an exci-tation-emission filter with an average wavelength of 495 mm
WHCO3 cells were seeded into 25 cm2 flasks as described
above. Cell cycle analyses were performed after 24 h ofexposure to 1 Â 10À6 M 2-ME at 378C. Cells weretrypsinized in equal volumes of trypsin (0.25%) and EDTA
(1 mM), fixed in 99.5% methanol and stored at À208C.
Data obtained from three independent experiments were
Methanol was removed by centrifugation at 200 Â g for
statistically analyzed for significance using the two-tailed
10 min. The sediments were resuspended in 1 ml 1% CaCl2
Student t-test for samples. Means are presented in bar charts,
and 50 mg mlÀ1 propidium iodide and incubated for 20 min
while shaking gently. Each analysis was based on at least
p-values < 0.05 were regarded statistically significant.
10 000 events employing a Coulter Epic-XS flow cytometer. The data were analyzed using a multicycle analysis program(MulticycleAV software).
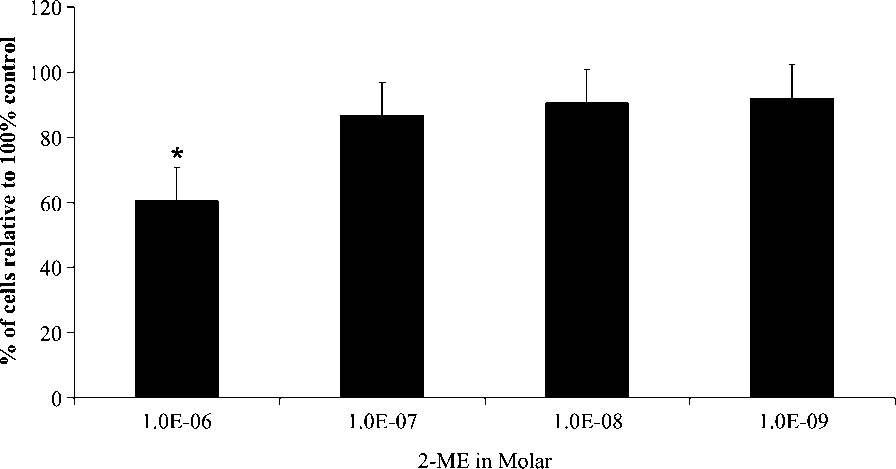
WHCO3 cell growth was expressed as a percentage of the
To visualize the effect of 2-ME on spindle formation in
control after exposure to different concentrations of 2-ME
WHCO3 cells, indirect immunofluorescence was employed.
(10À6, 10À7, 10À8, 10À9 M) for 72 h. 2-ME reduced cell
Cells (500 000) were seeded onto heat-sterilized glass cover
numbers to 60% when compared to vehicle-treated controls
slips in 6-well plates. After exposure to 2-ME or DMSO for
after exposure to 10À6 M 2-ME for 72 h. An à indicates a
24 h at 378C, cells were fixed in 10% formalin (2 mM EGTA
statistically significant p-value < 0.05 for growth inhibition
in PBS) for 10 min and permeabilized in ice-cold 97%
methanol containing 2 mM EGTA at À208C for 10 min. Subsequently cells were washed in PBS (3 Â 5 min) before
incubation for 1 h with a mouse monoclonal antibodyagainst human b-tubulin (Clone 2-28-33; 1:1000). After
Propidium iodide and hoechst 33342 staining. PI and HO
washing with PBS, cells were incubated with biotin-
staining were conducted to determine the presence of
conjugated anti-mouse IgG (Fab-specific, developed in
apoptotic cells after treatment with 2-ME. Viable and
goat) in FITC-conjugate diluent as secondary antibody for
apoptotic cells have intact cell membranes and are stained
Copyright # 2009 John Wiley & Sons, Ltd.
Cell Biochem Funct 2009; 27: 205–210.
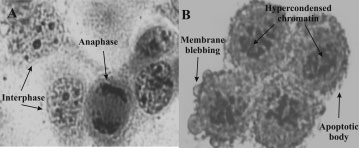
Hematoxylin and Eosin staining of WHCO3 control cells
exposed to 0.1% DMSO (vehicle) (A) and 1 Â 10À6 M 2-ME (B) for24 h (400 Â magnification). Clusters of rounded cells with hypercondensed
Dose-dependent study of WHCO3 cells treated with a dilution
chromatin, as well as apoptotic bodies are visible in the treated cells
series of 2-ME (10À6, 10À7, 10À8, 10À9 M) for 72 h. Cell numbers areexpressed as a percentage of the control. A 40% decrease in cell number wasnoted at 10À6 M of 2-ME WHCO3-treated cells. Ã indicates p-value < 0.05
intact nucleoli in contrast to cells treated with 2-ME thatshowed condensed chromatin, irregular nuclear membrane,
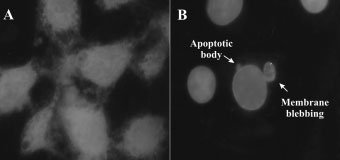
light blue, while cells that have lost their membrane integrity
and increased mitochondrial aggregation toward the nucleus
are stained bright red. After 2-ME treatment most cells
stained light blue (indicated in black and white) and wererounded in appearance due to a metaphase block and showed
apoptotic features including cytoplasmic shrinking, mem-brane blebbing, and apoptotic bodies (Figure 2A, B).
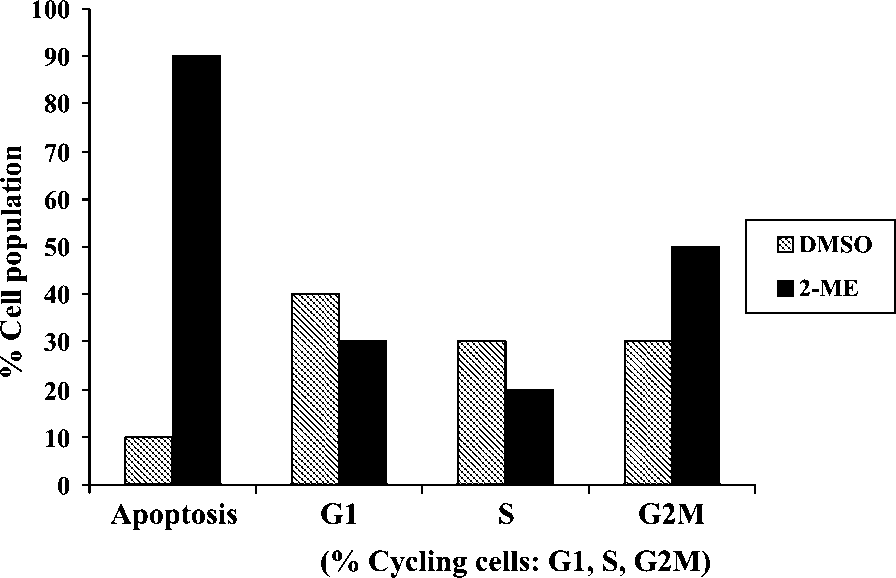
Quantitative analysis of DNA content was conducted bymeans of flow cytometry in order to determine the effects of2-ME on cycle progression after 24 h of exposure. 2-ME-
Hematoxylin and eosin staining (H and E staining)
treated cells revealed an increase in the sub G1/0 apoptotic
The antiproliferative effect of 2-ME observed above could
fraction when compared to vehicle-treated cells (Figure 5).
be attributed to either growth inhibition (cytostatic effect) or
An increase in the amount of cells in the G2/M-phase was
induction of cell death. Thus, morphological characteristics
of the cytoplasm and nuclear components of cells treatedwith 2-ME and DMSO respectively were studied by meansof hematoxylin and eosin staining to confirm 2-ME’santimitogenic effect (Figure 3A, B). After 2-ME treatment,most cells were rounded in appearance due to a metaphaseblock and showed apoptotic features including hypercon-densed chromatin, cytoplasmic shrinking, membrane bleb-bing, and apoptotic bodies when compared to their vehicle-treated controls (Figure 3A, B).
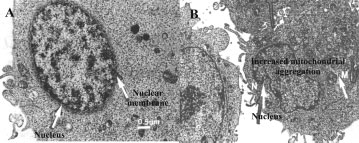
(A) Transmission electron microscopy of WHCO3 control cells
exposed to 0.1% DMSO (vehicle) (A) and 1 Â 10À6 M 2-ME (B) for 24 h.
Hypercondensed chromatin and increased mitochondrial aggregationaround the nucleus are visible (B). (Scale bar ¼ 0.5 mm)
TEM was employed to view subcellular structures in twodimensions. WHCO3 control cells (Figure 4A) revealed
Propidium iodide and Hoecsht 33342 staining of WHCO3 cells
exposed to 0.1% DMSO (vehicle) (A) and 1 Â 10À6 M 2-ME (B) for 24 h(black and white images; 400 Â magnification). 2-ME-treated cells are
The effect of 2-ME on cell cycle progression after 24 h of
rounded in appearance due to a metaphase block. Apoptotic features
exposure. 2-ME-treated cells presented with an increased sub G1/0 apoptotic
including cytoplasmic shrinking, membrane blebbing, and apoptotic bodies
fraction, as well as a G2/M-phase increase when compared to vehicle-treated
Copyright # 2009 John Wiley & Sons, Ltd.
Cell Biochem Funct 2009; 27: 205–210.
INFLUENCE OF 2-ME IN OESOPHAGEAL CARCINOMA CELLS
various cell lines that are affected by 2-ME at inhibitoryconcentrations ranging from 0.08 to 5 mM.
In the present study conducted, 2-ME was shown to exert
antiproliferative activity in the WHCO3 oesophagealcarcinoma cell line investigated. Morphological changesoccurring during apoptosis namely cell shrinkage, mem-brane blebbing condensation of nuclear chromatin intosharply delineated masses that become marginated againstthe nuclear membrane, as well as the formation of apoptoticbodies30 were demonstrated in 2-ME-treated WHCO3 cells.
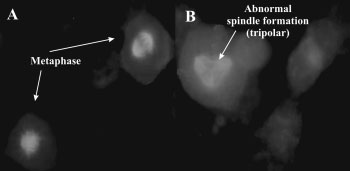
Immunofluorescent staining of b-tubulin in WHCO3 cells.
These data are consistent with previous results from our
2-ME-treated cells were rounded, accumulated in metaphase and spindle
laboratory where inhibition of cell growth in breast cancer
disruption with fragmented polar formations were evident when compared
cells (MCF-7) was demonstrated following 2ME treatment.
2ME-treated MCF-7 cells also exhibited abnormal meta-phase cells, membrane blebbing, apoptotic bodies, anddisturbed spindle formation. However, these observationswere either absent, or less pronounced in the non-tumorigenic MCF-12A cells.11
In this study, cells treated with 2-ME revealed an increase
in mitochondrial numbers aggregating around the nuclearenvelope. Mitochondria are important sensors and amplifiersin intracellular death signaling pathways and are corecomponents of the cell death machinery.14 Changes inmitochondrial membrane structure, either by disruption ofthe outer membrane or by Bax activation31 can lead toapoptosis. In addition, up-regulation of DR5 was also
Immunofluorescent detection of DR5 in 2-ME-treated WHCO3
demonstrated in 2-ME-treated cells and is consistent with
cells. Up-regulation of DR5 expression was observed as an increased
previous data where 2-ME was shown to up-regulate DR5
occurrence of white spots when compared to vehicle-treated controls
and sensitize cancer cells to TRAIL-induced apoptosis in
Furthermore, we have previously demonstrated a significant
increase in Cdc2 kinase activity in 2-ME-treated cells whencompared to vehicle-treated controls in WHCO3 cells.10 Cdc2
Since previous research has shown that 2-ME induces cell
kinase activity was statistically significantly increased (1.7-
death by causing microtubule disruption and blocking cells
fold) ( p < 0.005) after 2-ME exposure when compared to
in metaphase in other cell lines, the influence of 2-ME was
vehicle-treated controls. Our observation contributes to the
subsequently investigated on spindle formation in WHCO3
elucidating of the mechanism of action in WHCO3
cells by means of immunofluorescent staining of b-tubulin.
oesophageal carcinoma cells and reveals that 2-ME causes
2-ME-treated cells were rounded, accumulated in metaphase
a metaphase arrest, disrupts mitotic spindle formation,
and also showed spindle disruption with fragmented polar
enhances Cdc2 kinase activity leading to persistence of the
formations when compared to vehicle-treated controls
spindle checkpoint, and thus prolonged metaphase arrest
culminating in the induction of apoptosis in WHCO3 cells. The observed up-regulation of DR5 further supports theproposed mechanism of apoptosis induction via the extrinsic
pathway in WHCO3 oesophageal carcinoma cells.
To investigate whether the extrinsic pathway of apoptosiswas activated after treatment with 2-ME, DR5 was chosen asa marker. Immunofluorescent detection of DR5 in 2-ME-
treated WHCO3 cells demonstrated an up-regulation of DR5expression when compared to vehicle-treated controls
This research was supported by grants from the Medical
Research Council of South Africa (AG374, AK076), theCancer Association of South Africa (AK246), and theStruwig-Germeshuysen Cancer Research Trust of South
Africa (AJ038). Electron microscopy was conducted at
Previous research has revealed that 2-ME plays an important
the Electron microscopy Unit at the University of Pretoria
role in the induction of apoptosis and especially in actively
and flow cytometric analysis was performed at the Depart-
proliferating cancerous cells.3,5,16,22,28,29 Pribluda et al.16
ment of Pharmacology at the Faculty of Health Sciences
accounted of the antiproliferative effects of 2-ME by listing
Copyright # 2009 John Wiley & Sons, Ltd.
Cell Biochem Funct 2009; 27: 205–210.
17. Banerjee SK, Zoubine MN, Sarkar DK, Weston AP, Shah JH, Campbell
DR. 2-Methoxyestradiol blocks estrogen-induced rat pituitary tumor
1. Dingli D, Timm M, Russell SJ, Witzig TE, Rajkumar SV. Promising
growth and tumor angiogenesis: possible role of vascular endothelial
preclinical activity of 2-methoxyestradiol in multiple myeloma. Clin
growth factor. Anticancer Res 2000; 20: 2641–2645.
18. Lippert TH, Adlercreutz H, Berger MR, Seeger H, Elger W, Mueck AO.
2. Lottering ML, Haag M, Seegers JC. Effects of 17 beta-estradiol
Effect of 2-methoxyestradiol on the growth of methyl-nitroso-urea
metabolites on cell cycle events in MCF-7 cells. Cancer Res 1992;
(MNU)-induced rat mammary carcinoma. J Steroid Biochem Mol Biol
3. Lis A, Ciesielski MJ, Barone TA, Scott BE, Fenstermaker RA, Plunkett
19. Lakhani NJ, Sarkar MA, Venitz J, Figg WD. 2-Methoxyestradiol,
RJ. 2-Methoxyestradiol inhibits proliferation of normal and neoplastic
a promising anticancer agent. Pharmacotherapy 2003; 23: 165–
glial cells, and induces cell death, in vitro. Cancer Lett 2004; 213: 57–65.
4. D’Amato RJ, Lin CM, Flynn E, Folkman J, Hamel E. 2-Methoxyes-
20. Basu A, Castle VP, Bouziane M, Bhalla K, Haldar S. Crosstalk between
tradiol, an endogenous mammalian metabolite, inhibits tubulin
extrinsic and intrinsic cell death pathways in pancreatic cancer: syner-
polymerization by interacting at the colchicine site. Proc Natl Acad
gistic action of estrogen metabolite and ligands of death receptor
family. Cancer Res 2006; 66: 4309–4318.
5. Mooberry SL. Mechanism of action of 2-methoxyestradiol: new devel-
21. Golab J, Nowis D, Skrzycki M, et al. Antitumor effects of photo-
opments. Drug Resist Updat 2003; 6: 355–361.
dynamic therapy are potentiated by 2-methoxyestradiol. A superoxide
6. Wang SH, Myc A, Koenig RJ, Bretz JD, Arscott PL, Baker JR.
dismutase inhibitor. J Biol Chem 2003; 278: 407–414.
2-Methoxyestradiol, an endogenous estrogen metabolite, induces
22. Fotsis T, Zhang Y, Pepper MS, et al. The endogenous oestrogen
thyroid cell apoptosis. Mol Cell Endocrinol 2000; 165: 163–172.
metabolite 2-methoxyoestradiol inhibits angiogenesis and suppresses
7. Nakagawa-Yagi Y, Ogane N, Inoki Y, Kitoh N. The endogenous
tumour growth. Nature 1994; 368: 237–239.
estrogen metabolite 2-methoxyestradiol induces apoptotic neuronal
23. Seegers JC, Aveling ML, Van Aswegen CH, Cross M, Koch F, Joubert
cell death in vitro. Life Sci 1996; 58: 1461–1467.
WS. The cytotoxic effects of estradiol-17 beta, catecholestradiols and
8. Joubert A, Maritz C, Joubert F. Bax/Bcl-2 expression levels of
methoxyestradiols on dividing MCF-7 and HeLa cells. J Steroid
2-methoxyestradiol-exposed esophageal cancer cells. Biomed Res
24. Maran A, Zhang M, Kennedy AM, et al. 2-methoxyestradiol induces
9. Joubert A, Maritz C, Joubert F. Influence of prostaglandin A2 and
interferon gene expression and apoptosis in osteosarcoma cells. Bone
2-methoxyestradiol on Bax and Bcl-2 expression levels in cervical
carcinoma cells. Biomed Res 2005; 26: 87–90.
25. Gillies RJ, Didier N, Denton M. Determination of cell number in
10. Joubert A, Marais S. Influence of 2-methoxyestradiol on cell
monolayer cultures. Anal Biochem 1986; 159: 109–113.
morphology and cdc2 kinase activity in WHCO3 esophageal carcinoma
26. Ciancio G, Pollack A, Taupier MA, Block NL, Irvin GL III. Measure-
ment of cell-cycle phase-specific cell death using Hoechst 33342 and
11. Van ZC Lottering, Steffens ML, Joubert FA. In vitro effects of
propidium iodide: preservation by ethanol fixation. J Histochem Cyto-
2-methoxyestradiol on MCF-12A and MCF-7 cell growth, morphology
and mitotic spindle formation. Cell Biochem Funct 2008; 26: 632–642.
27. Huppertz B, Frank HG, Kaufmann P. The apoptosis cascade–morpho-
12. Andreassen PR, Margolis RL. Microtubule dependency of p34cdc2
logical and immunohistochemical methods for its visualization. Anat
inactivation and mitotic exit in mammalian cells. J Cell Biol 1994; 127:
28. Amant F, Lottering ML, Joubert A, Thaver V, Vergote I, Lindeque BG.
13. LaVallee TM, Zhan XH, Johnson MS, et al. 2-methoxyestradiol up-
2-Methoxyestradiol strongly inhibits human uterine sarcomatous cell
regulates death receptor 5 and induces apoptosis through activation of
growth. Gynecol Oncol 2003; 91: 299–308.
the extrinsic pathway. Cancer Res 2003; 63: 468–475.
29. Perez-Stable C. 2-Methoxyestradiol and paclitaxel have similar effects
14. Qanungo S, Basu A, Das M, Haldar S. 2-Methoxyestradiol induces
on the cell cycle and induction of apoptosis in prostate cancer cells.
mitochondria dependent apoptotic signaling in pancreatic cancer cells.
30. Saraste A, Pulkki K. Morphologic and biochemical hallmarks of
15. Thaver V. Mechanism(s) of apoptosis induction by 2-methoxyestradiol,
apoptosis. Cardiovasc Res 2000; 45: 528–537.
an estrogen metabolite, in SNO and WHCO3 oesophageal carcinoma
31. Burns TF, El-Deiry WS. The p53 pathway and apoptosis. J Cell Physiol
cell lines. University of Pretoria. 2003.
16. Pribluda VS, Gubish ER Jr, LaVallee TM, Treston A, Swartz GM,
32. Ibrahim SM, Ringel J, Schmidt C, et al. Pancreatic adenocarcinoma cell
Green SJ. 2-Methoxyestradiol: an endogenous antiangiogenic and
lines show variable susceptibility to TRAIL-mediated cell death.
antiproliferative drug candidate. Cancer Metastasis Rev 2000; 19:
Copyright # 2009 John Wiley & Sons, Ltd.
Cell Biochem Funct 2009; 27: 205–210.
Gynaecologie Miskraam / vaginaal bloedverlies Bron: teksten zijn grotendeels overgenomen van NVOG 2005 Wat betekent vaginaal bloedverlies in het begin van de zwangerschap? Bloedverlies in de eerste maanden van de zwangerschap is meestal onschuldig en kan verschillende oorzaken hebben. Veelal is het bloedverlies onschuldig en stopt vanzelf. In de helft van het aantal keren
05.31.06; Revised 06.12.06; 07.24.06; 12.30.06, 07.07,09.07,04.08 Items in bold italics are recent changes/additions Adults are age 18 years and older; children are greater than 6 months and less than 18 years of age; neonates or infants are less than 6 months of age. Provide only the first initial of the last name. Provide only the first initial of the first name. This is optional; for
 Hematoxylin and Eosin staining of WHCO3 control cells
exposed to 0.1% DMSO (vehicle) (A) and 1 Â 10À6 M 2-ME (B) for24 h (400 Â magnification). Clusters of rounded cells with hypercondensed
Dose-dependent study of WHCO3 cells treated with a dilution
chromatin, as well as apoptotic bodies are visible in the treated cells
series of 2-ME (10À6, 10À7, 10À8, 10À9 M) for 72 h. Cell numbers areexpressed as a percentage of the control. A 40% decrease in cell number wasnoted at 10À6 M of 2-ME WHCO3-treated cells. Ã indicates p-value < 0.05
intact nucleoli in contrast to cells treated with 2-ME thatshowed condensed chromatin, irregular nuclear membrane,
light blue, while cells that have lost their membrane integrity
and increased mitochondrial aggregation toward the nucleus
are stained bright red. After 2-ME treatment most cells
stained light blue (indicated in black and white) and wererounded in appearance due to a metaphase block and showed
apoptotic features including cytoplasmic shrinking, mem-brane blebbing, and apoptotic bodies (Figure 2A, B).
Hematoxylin and Eosin staining of WHCO3 control cells
exposed to 0.1% DMSO (vehicle) (A) and 1 Â 10À6 M 2-ME (B) for24 h (400 Â magnification). Clusters of rounded cells with hypercondensed
Dose-dependent study of WHCO3 cells treated with a dilution
chromatin, as well as apoptotic bodies are visible in the treated cells
series of 2-ME (10À6, 10À7, 10À8, 10À9 M) for 72 h. Cell numbers areexpressed as a percentage of the control. A 40% decrease in cell number wasnoted at 10À6 M of 2-ME WHCO3-treated cells. Ã indicates p-value < 0.05
intact nucleoli in contrast to cells treated with 2-ME thatshowed condensed chromatin, irregular nuclear membrane,
light blue, while cells that have lost their membrane integrity
and increased mitochondrial aggregation toward the nucleus
are stained bright red. After 2-ME treatment most cells
stained light blue (indicated in black and white) and wererounded in appearance due to a metaphase block and showed
apoptotic features including cytoplasmic shrinking, mem-brane blebbing, and apoptotic bodies (Figure 2A, B).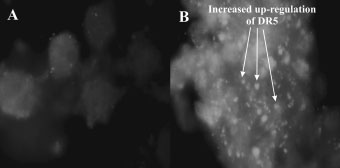
 INFLUENCE OF 2-ME IN OESOPHAGEAL CARCINOMA CELLS
various cell lines that are affected by 2-ME at inhibitoryconcentrations ranging from 0.08 to 5 mM.
INFLUENCE OF 2-ME IN OESOPHAGEAL CARCINOMA CELLS
various cell lines that are affected by 2-ME at inhibitoryconcentrations ranging from 0.08 to 5 mM.