Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Fayoum.edu.eg
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Mar. 2003, p. 1115–1119
0066-4804/03/$08.00ϩ0 DOI: 10.1128/AAC.47.3.1115–1119.2003
Copyright 2003, American Society for Microbiology. All Rights Reserved.
Mercury Resistance Determinants Related to Tn21, Tn1696, and
Tn5053 in Enterobacteria from the Preantibiotic Era
Ashraf M. M. Essa, Daniel J. Julian, Stephen P. Kidd, Nigel L. Brown, and Jon L. Hobman*
School of Biosciences, The University of Birmingham, Edgbaston, Birmingham B15 2TT, United Kingdom
Received 9 July 2002/Returned for modification 4 October 2002/Accepted 2 December 2002
Three mer transposons from the Murray collection of preantibiotic enterobacteria show >99% sequence identity to current isolates. Tn5073 is most closely related to Tn5036 and Tn1696, and Tn5074 is most closely related to Tn5053. Tn5075 is most closely related to Tn21 but lacks integron In2 and is flanked by insertion elements.
Tn21 encodes mercuric ion resistance (Hgr) and contains the
PCR was performed with part of the mer operon from each
class I integron In2, encoding resistance to sulfonamides (sul)
plasmid as described elsewhere (3). PCR products purified by
and streptomycin-spectinomycin (aadA) (9, 18). It is carried by
using a QIAquick PCR purification kit (Qiagen, Ltd., Crawley,
the conjugative plasmid NR1 (R100), which was isolated in
United Kingdom) were sequenced with the Big Dye terminator
Japan in the 1950s (21). More recently, Tn21 (18) and other
cycle sequencing kit (PE Applied Biosystems, Warrington,
Tn21-like transposons carrying integron-associated antibiotic
United Kingdom) and an Applied Biosystems 3700 sequencer,
resistance (7) have been detected in Escherichia coli from ag-
according to the manufacturer’s protocols. Further sequence
ricultural (1, 28) and nonclinical sources (19), as well as from
analysis was performed by using primers designed from the
mercury amalgam-exposed, gram-negative gut bacteria (16, 17,
sequences so obtained and from merA gene primers (5) (the
33), clinical bacterial isolates (13, 31, 36), and intercontinental
primers used are described at http://www.biosciences.bham.ac
plasmids carried by clinical isolates (10, 22).
.uk/labs/brown/mer_primers.htm). The transposon terminal in-
It is now thought that Tn21 evolved by the insertion of an
verted repeat DNA and flanking sequences were amplified by
In2 ancestor (lacking IS1353) into the urf2M gene of a hypo-
inverse PCR (26). Genetic maps of the sequenced mer operons
thetical mercury resistance transposon, Tn21⌬ (18) (also called
are shown in Fig. 1A. DNA alignments and analysis were
TnX [25]), probably catalyzed by transposition proteins en-
performed with the University of Wisconsin Genetics Com-
coded in trans (2, 18). A similar event led to the formation of
puter Group version 9.0 suite of programs at the University of
Tn1696 from plasmid R1033, where In4 inserted at the res site
of a Tn5036-like mer transposon (25).
The three mer operons that we sequenced represent differ-
Hughes and Datta identified three Hgr bacterial strains,
ent lineages and are not closely related to each other, but they
M426, M567, and M634, from a total of 433 strains from the
are closely related to mer transposons isolated since the 1950s.
Murray collection of preantibiotic era enterobacteria (11). In
Table 2 shows the percent identities between the sequences of
this study, we sequenced the mer operons from M426, M567,
the genes from Tn5073 (Klebsiella pneumoniae M426), Tn5074
and M643 in order to investigate the relationships between Hgr
(Morganella morganii M567), Tn5075 (E. coli M634) and pub-
sequences from clinical bacteria that had been isolated before
antibiotics came into widespread use and present-day Hgr se-
The sequenced merRTPCAD genes (3,788 bp) of the Tn5073mer operon (Fig. 1A) had the highest identity at the DNA level
The plasmids, bacterial strains, antimicrobial resistance of
to those from Tn5036 (35), a Tn5036-like mer transposon from
these strains, and the 16S ribosomal DNA sequence identifi-
Salmonella enterica serovar Typhi CT18 plasmid pHCM1 (24),
cations of the three Hgr strains from this study (determined as
and to the sequenced merAD genes of Tn1696, which carries
previously described [32]) are shown in Table 1. All bacteria
In4 (25) (Table 2). In comparison to the sequence encoded by
were grown at 37°C in Luria broth (LB) or on LB agar (27).
Tn5036, there were two amino acid differences in the sequence
Hgr plasmids from M426, M567, and M634 were mated with E.
encoded by Tn5073: MerR A173V and MerA V2503A. The
coli TG2 (20), and Hgr transconjugants were grown overnight
Tn5073 merT gene carries five GTCTGAACCACAAAA du-
on LB agar plates containing tetracycline (15 g/ml) and
plications at the 5Ј end (Fig. 1A). Multiple repeats of this
sequence have also been observed in enterobacterial mercury
The E. coli TG2 Hgr transconjugants from each of the three
resistance determinants from primates (16) and in Tn5036,
Murray strains contained an ϳ60-kb plasmid that conferred
Hgr. Plasmid DNA was isolated by standard methods (27), and
The sequenced merRTPFADE genes (3,647 bp) from the
Tn5074 mer operon (Fig. 1A) had the highest identity at the
DNA level to those from Tn5053 (14) and pMER327/419 (8)
* Corresponding author. Mailing address: School of Biosciences,
(Table 2). In comparison to the sequence encoded by Tn5053,
The University of Birmingham, Edgbaston, Birmingham B15 2TT,
United Kingdom. Phone: 44 (0)121 414 6558. Fax: 44 (0)121 414 5925.
there were four amino acid differences in the sequence en-
coded by Tn5074: MerR A1193S and K1213Q and MerA
TABLE 1. Bacterial strains and plasmids, antibiotic resistance phenotypes, 16S rRNA sequences, and sources of the strains used in this study
K-12 lac-pro⌬srl-recA 306::Tn10proAϩ Bϩ laclq lacZ⌬M15]
a Tcr, tetracycline resistance; Rifr, rifampicin resistance; Apr, ampicillin resistance; Kmr, kanamycin resistance. b NCTC, National Collection of Type Cultures, Colindale, London, United Kingdom.
V2323A and S2893G. The Tn5053 type mer operon was first
Tn21, resulting in a frameshift and a 314-amino-acid protein,
isolated in environmental bacteria (8, 14) and has also been
rather than the 328-amino-acid protein predicted previously
found in the fecal flora of primates and humans exposed to
The insertion (IS) elements IS5075L and IS5075R (which
In total, 11,298 bp of Tn5075 were sequenced. The 3,962-bp
are 1,351 bp and Ͼ99.5% identical to each other) flank
mer operon from Tn5075 carrying merRTPCADE has the high-
Tn5075, forming a composite transposon (Fig. 1A). IS5075L
est identity at the DNA level to the equivalent regions of Tn21
and IS5075R belong to the IS110 family and are Ͼ99.6%
(18) and the Tn21-like mer transposon from S. enterica serovar
identical to the IS elements flanking the Tn21-like mer trans-
Typhi CT18 plasmid pHCM1 (24) (Fig. 1A) (Table 2). The
poson from plasmid pHCM1 (Fig. 1A). IS5075L and IS5075R
Tn5075 res site and the transposition genes (3,529 bp) tnpR and
are between 91 and 93% identical to IS element sequences
tnpA again had the highest identity to those of Tn21. In com-
flanking other mercury resistance-encoding genes from gram-
parison to the sequence encoded by Tn21, there were three
amino acid differences in the sequence encoded by Tn5075:
MerA, Q5583H; TnpR, T1653A; and TnpA R4553A. Most
Transposition of the Hgr phenotype was determined by a
importantly, Tn5075 did not carry the integron In2, which in
mate-out assay (15) with E. coli TG2 carrying plasmid RK2,
Tn21 is located between urf2 and the putative transposition
conjugated with Hgr plasmids from M426, M567, or M634, into
gene tnpM (12) (Fig. 1A). Instead, in Tn5075, there is a single
E. coli KH802. Transconjugants were selected on LB agar
945-bp open reading frame (urf2M) of unknown function
plates containing HgCl2 (20 g/ml) and carbenicillin (200 g/
rather than the predicted 987-bp hypothetical urf2M from
ml), and the donor strain was counterselected with rifampin
Tn21⌬ proposed by Liebert et al. (18). Tn5075 urf2M is 97.1%
(50 g/ml). Purified plasmid RK2 DNA from the transconju-
identical at the DNA level to the hypothetical urf2M that
gants was analyzed for transposon insertion by PstI digestion
Liebert et al. (18) postulated to exist in the ancestor of Tn21.
and gel electrophoresis (27). Tn5073 transposed into plasmid
The reason for this lower percent identity is a 23-bp deletion in
RK2 at a frequency of 6.3 ϫ 10Ϫ5 per donor cell. Tn5074
the Tn5075 urf2M sequence compared to the tnpM sequence in
transposed into RK2 at a frequency of 2.9 ϫ 10Ϫ4 per donor
FIG. 1. (A) Genetic maps of the sequenced regions of Tn5073, Tn5074, Tn5075, Tn21 (18), and the Tn21-like transposon from pHCM1 (24).
Vertical black bars represent the 10- to 12-bp inverted repeat sequences flanking the IS elements. Vertical unfilled bars represent the 38bp inverted
repeat sequences at the ends of the class II transposition module. The transposition (tnp) regions consist of the transposase gene (tnpA), the
resolvase gene (tnpR), the putative transposon regulator (tnpM), and the resolution site, res. urf2M in Tn5075 is of unknown function. The points
at which In2 or antibiotic resistance-encoding IS elements have inserted into Tn21 and pHCM1 are marked with black arrows. The mercury
resistance-encoding operons consist of the regulatory genes merR and merD, the mercury transport genes merT, merP, and merC or merF, the
putative transporter merE, and the gene encoding mercuric reductase, merA. urfI and urf2 are of unknown function. All mer genes are marked in
the figure with a single letter. The position of the multiple DNA sequence repeats found in merT from Tn5073 are indicated (). (B) Postulated
evolutionary pathway for Tn5075, Tn21, and the Tn21-like transposon in pHCM1. An ancestral mer transposon (a) could acquire either IS5075L
and IS5075R to become Tn5075 (b) or an integron related to In2, resulting in the formation of Tn21 (c). Gene insertions and deletions in In2 would
lead to Tn21 variants (d), or Tn21 could have acquired IS elements, leading to the formation of a precursor to the Tn21-like transposon in pHCM1
(e). Deletions of transposition genes and partial deletion of In2, followed by insertion of antibiotic resistance gene-carrying IS elements, would
result in the formation of the Tn21-like transposon in pHCM1 (f).
TABLE 2. Comparison of DNA sequence identities between Tn5073, Tn5074, Tn5075, and
published DNA sequences from other transposons
% Identity to mer transposon (genes)a
Ͼ99.9 Tn5036-like transposon pHCM1
Ͼ99.9 Tn21-like transposon from pHCM1 (merRTPCADE)
Ͼ99.8 Tn21 (res tnpA tnpR)
Ͼ97.1 hypothetical urf2m from Tn21⌬
Ͼ99.6 Tn21-like transposon from pHCM1 (IS elements)
Ͼ91.0 IS4321L, IS4321R, from R751
cell. We found no transposition of Tn5075 in our assays, which
are as follows: Tn5073 (strain M426), AF461013; Tn5074 (strain
could detect frequencies of Ͼ10Ϫ7.
M567), AF461012; and Tn5075 (strain M634), AF457211. The
In conclusion, the internal genetic structure of Tn5075 is
16S rRNA gene sequence accession number for strain M567 is
consistent with the recently proposed structure of Tn21⌬
(Fig. 1B) (2, 18), and Tn5073 is closely related to Tn5036
and Tn1696 (25). The sequence data from Tn5075 support
We thank Julian Davies, Didier Mazel, and Barry Holmes for their
the hypothesis that Tn21 evolved from an Hgr transposon sim-
help. We also thank Chris Thomas and Peter Strike for the gifts of
ilar to Tn5075, rather than from Tn2613 (29). Tn1696 and Tn21
plasmids and strains and Gennady Kholodii and anonymous referees
for suggested improvements to the paper.
represent independent lineages of mer transposons that have
This work was supported by a postgraduate research scholarship to
acquired integrons (25). The dates of isolation of Tn5073
A.M.E. from the Egyptian Ministry of Higher Education, and the work
(1940) and Tn5075 (1931) and the close relationship of these
was partially supported by a BBSRC grant 6/G07943 to N.L.B. and a
transposons to other lineages (Tn5073 merAD genes are Ͼ99.9%
BBSRC/Wellcome Trust Joint Infrastructure Fund Grant (6/JIF13209).
identical to those of Tn1696; Tn5075 is Ͼ99.6% identical to
Support in bioinformatics was from MRC grant G.4600017.
Tn21 except where In2 is not present) are consistent with the
idea that integrons transposed into preexisting clinical Hgr
REFERENCES
transposons. The Tn5074 mer operon, isolated from a clini-
1. Bass, L., C. A. Liebert, M. D. Lee, A. O. Summers, D. G. White, S. G. Thayer,
cal source, has the greatest DNA identity to the Tn5053 and
and J. G. Maurer. 1999. Incidence and characterization of integrons, genetic
elements mediating multiple-drug resistance, in avian Escherichia coli. An-
pMER327/419 mer operons, which have been isolated from
timicrob. Agents Chemother. 43:2925–2929.
both environmental and nonclinical (16, 20) sources.
2. Brown, H. J., H. W. Stokes, and R. M. Hall. 1996. The integrons In0, In2, and
In5 are defective transposon derivatives. J. Bacteriol. 178:4429–4437.
The DNA sequence data suggest that Tn5075, the Tn21-like
3. Bruce, K. D., A. M. Osborn, A. J. Pearson, P. Strike, and D. A. Ritchie. 1995. mer transposon in pHCM1, and Tn21 had a common ancestor
Genetic diversity within mer genes directly amplified from communities of
and may have evolved as shown in Fig. 1B; i.e., an ancestral
noncultivated soil and sediment bacteria. Mol. Ecol. 4:605–612.
4. Gibson, T. 1984. Studies on the Epstein-Barr virus genome. Ph.D. thesis. mer transposon acquired IS5075L and IS5075R, leading to the
University of Cambridge, Cambridge, United Kingdom.
formation of Tn5075. Alternatively, an integron related to In2
5. Glendinning, K. G. 2000. Studies on mercuric reductase and thermophilic
inserted into the ancestral mer transposon, leading to the for-
mercury resistance. Ph.D. thesis. The University of Birmingham, Birming-
mation of Tn21. Gene insertions and deletions within the in-
6. Griffin, H. G., T. J. Foster, S. Silver, and T. K. Misra. 1987. Cloning and
tegron in Tn21 could lead to the formation of Tn21 variants
DNA sequence of mercuric and organomercurial-R determinants of plasmid
(7). Acquisition of IS5075-like elements by Tn21 could have
pDU1358. Proc. Natl. Acad. Sci. USA 84:3112–3116.
7. Grinsted, J., F. de la Cruz, and R. Schmitt. 1990. The Tn21 subgroup of
led to the formation of a hypothetical mer transposon. Dele-
bacterial transposable elements. Plasmid 26:163–189.
tion of res and tnpR and partial deletion of In2 and tnpA from
8. Hobman, J., G. Kholodii, V. Nikiforov, D. A. Ritchie, P. Strike, and O.
this transposon followed by insertion of antibiotic resistance-
Yurieva. 1994. The sequence of the mer operon of pMER327/419 and trans-
poson ends of pMER327/419, 330, and 05. Gene 277:73–78.
carrying IS elements, could have led to the formation of the
9. Hobman, J. L., and N. L. Brown. 1997. Bacterial mercury-resistance genes,
Tn21-like transposon in pHCM1 (24). Although the Tn21-like
p. 527–568. In A. Sigel and H. Sigel (ed.), Metal ions in biological systems.
transposon in pHCM1 is flanked by IS elements which are
10. Hopkins, J. D., T. F. O’Brien, and M. Syvanen. 1988. Functional and struc-
99.6% identical to IS5075L and IS5075R from Tn5075, it is
tural map of pLST1000: a multiresistance plasmid widely distributed in
more closely related to Tn21 than to Tn5075 because it con-
Enterobacteriaceae. Plasmid 20:163–166.
11. Hughes, V. M., and N. Datta. 1983. Conjugative plasmids in bacteria of the
tains a vestige of the In2 sequence, which Tn5075 does not and
Јpre-antibiotic’ era. Nature 302:725–726.
is 100% identical to Tn21 across the mer genes and tnpA.
12. Hyde, D. R., and C. P. D. Tu. 1985. tnpM: a novel regulatory gene that Nucleotide sequence accession numbers. The GenBank ac-
enhances Tn21 transposition and suppresses cointegrate resolution. Cell
42:629–638.
cession numbers of the mer sequences determined in this study
13. Jones, M. E., E. Peters, A.-M. Weersink, A. Fluit, and J. Verhoef. 1997.
Widespread occurrence of integrons causing multiple antibiotic resistance in
Complete genome sequence of a multiple drug resistant Salmonella enterica
bacteria. Lancet 349:1742–1743.
serovar Typhi CT18. Nature 413:848–852.
14. Kholodii, G. Y., S. Z. Mindlin, I. A. Bass, O. V. Yurieva, S. V. Minakhina,
25. Partridge, S. R., H. J. Brown, H. W. Stokes, and R. M. Hall. 2001. Trans- and V. G. Nikiforov. 1995. Four genes, two ends, and a res region are
posons Tn1696 and Tn21 and their integrons In4 and In2 have independent
involved in the transposition of Tn5053: a paradigm for a novel family of
origins. Antimicrob. Agents Chemother. 45:1263–1270.
transposons carrying either a mer operon or an integron. Mol. Microbiol.
26. Riley, J., R. Butler, R. Finniear, D. Jenner, S. Powell, R. Anand, J. C. Smith, 17:1189–1200. and A. F. Markham. 1990. A novel, rapid method for the isolation of
15. Kholodii, G., O. Yurieva, S. Mindlin, Z. Gorlenko, V. Rybochkin, and V.
terminal sequences from yeast artificial chromosome (YAC) clones. Nucleic
Nikiforov. 2000. Tn5044, a novel Tn3 family transposon coding for temper-
Acids Res. 18:2887–2890.
ature-sensitive mercury resistance. Res. Microbiol. 151:291–312.
27. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a
16. Liebert, C. A., J. Wireman, T. Smith, and A. O. Summers. 1997. Phylogeny
laboratory manual, 2nd ed., vol 3. Cold Spring Harbor Laboratory Press,
of mercury resistance (mer) operons of gram-negative bacteria isolated from
the fecal flora of primates. Appl. Environ. Microbiol. 63:1066–1076.
28. Sunde, M., and H. Sørum. 2001. Self-transmissible multidrug resistance
17. Liebert, C. A., J. Wireman, T. Smith, and A. O. Summers. 1997. The impact
plasmids in Escherichia coli of the normal intestinal flora of healthy swine.
of mercury released from dental “silver” fillings on antibiotic resistances in
Microb. Drug Resist. 7:191–196.
the primate oral and intestinal bacterial flora, p. 441–460. In A. Sigel and H.
29. Tanaka, M., T. Yamamoto, and T. Sawai. 1983. Evolution of complex resis-
Sigel (ed.), Metal ions in biological systems. Marcel Dekker, Inc., New York,
tance transposons from an ancestral mercury transposon. J. Bacteriol. 153:
18. Liebert, C. A., R. M. Hall, and A. O. Summers. 1999. Transposon Tn21,
30. Thorsted, P. B., D. P. Macartney, P. Akhtar, A. S. Haines, N. Ali, P. David-
flagship of the floating genome. Microbiol. Mol. Biol. Rev. 63:507–522. son, T. Stafford, M. J. Pocklington, W. Pansegrau, B. M. Wilkins, E. Lanka,
19. Mazel, D., B. Dychinco, V. A. Webb, and J. Davies. 2000. Antibiotic resis- and C. M. Thomas. 1998. Complete sequence of the IncP plasmid R751:
tance in the ECOR collection: integrons and identification of a novel aad
implications for evolution and organisation of the IncP backbone. J. Mol.
gene. Antimicrob. Agents Chemother. 44:1568–1574.
Biol. 282:969–990.
20. Mindlin, S., G. Kholodii, Z. Gorlenko, S. Minakhina, L. Minakhin, E.
31. White, P. A., C. J. McIver, and W. D. Rawlinson. 2001. Integrons and gene Kalyaeva, A. Kopteva, M. A. Petrova, O. V. Yurieva, and V. Nikiforov. 2001.
cassettes in the Enterobacteriaceae. Antimicrob. Agents Chemother. 45:
Mercury resistance transposons of gram-negative bacteria, and their classi-
fication. Res. Microbiol. 152:811–822.
32. Wintzingerode, F. V., U. B. Gobel, and E. Stackebrandt. 1997. Determination
21. Nakaya, R., A. Nakamura, and Y. Murata. 1960. Resistance transfer agents
of microbial diversity in environmental samples: pitfalls of PCR-based rRNA
in Shigella. Biochem. Biophys. Res. Commun. 3:654–659.
analysis. FEMS Microbiol. Rev. 21:213–229.
22. O’Brien, T. F., M. del Pilar Pla, K. H. Mayer, H. Kishi, E. Gilleece, M.
33. Wireman, J., C. A. Liebert, T. Smith, and A. O. Summers. 1997. Association Syvanen, and J. D. Hopkins. 1985. Intercontinental spread of a new antibi-
of mercury resistance with antibiotic resistance in the gram-negative fecal
otic resistance gene on an epidemic plasmid. Science 230:87–88.
bacteria of primates. Appl. Environ. Microbiol. 63:4494–4503.
23. Ogawa, H. I., C. L. Tolle, and A. O. Summers. 1984. Physical and genetic
34. Wood, W. B. 1966. Host specificity of DNA produced by Escherichia coli:
map of the organomercury resistance (Omr) and inorganic mercury resis-
bacterial mutations affecting the restriction and modification of DNA. J.
tance (Hgr) loci of the IncM plasmid R831b. Gene 32:311–320.
Mol. Biol. 16:118–133.
24. Parkhill, J., G. Dougan, K. D. James, N. R. Thomson, D. Pickard, J. Wain,
35. Yurieva, O., G. Kholodii, L. Minakhin, Z. Gorlenko, E. Kalyaeva, S. Mind- C. Churcher, K. L. Mungall, S. D. Bentley, M. T. G. Holden, M. Sebaihia, S. lin, and V. Nikiforov. 1997. Intercontinental spread of promiscuous mercury Baker, D. Basham, K. Brooks, T. Chillingworth, P. Connerton, A. Cronin, P.
resistance transposons in environmental bacteria. Mol. Microbiol. 24:321– Davis, R. M. Davies, L. Dowd, N. White, J. Farrar, T. Feltwell, N. Hamlin, A. Haque, T. T. Hien, S. Holroyd, K. Jagels, A. Krogh, T. S. Larsen, S.
36. Zu¨hlsdorf, M. T., and B. Weidemann. 1992. Tn21-specific structures in gram- Leather, S. Moule, P. O ´ ’Gaora, C. Parry, M. Quail, K. Rutherford, M.
negative bacteria from clinical isolates. Antimicrob. Agents Chemother. 36: Simmonds, J. Skelton, K. Stevens, S. Whitehead, and B. G. Barrell. 2001.
Summary of Hall, et al. study “Mid-Century Warming in the Los Angeles Region.” Prepared by Paul Bunje PhD The first results from the “Climate Change in the Los Angeles Region” project, sponsored by the City of Los Angeles and US Department of Energy covers temperature changes. Future results will cover other elements of climate change (precipitation, snowpack, winds, clouds, etc.). T
TO UEFA MEMBER ASSOCIATIONS TO CLUBS PARTICIPATING IN UEFA COMPETITIONS For the attention of the President and the General Secretary 2011 List of Prohibited Substances In accordance with paragraph 4.01 of the 2010 UEFA Anti-Doping Regulations, the 2011 WADA Prohibited List applies to all UEFA competitions, with effect from 1 January 2011 . For this purpose, we enclose the new list of p
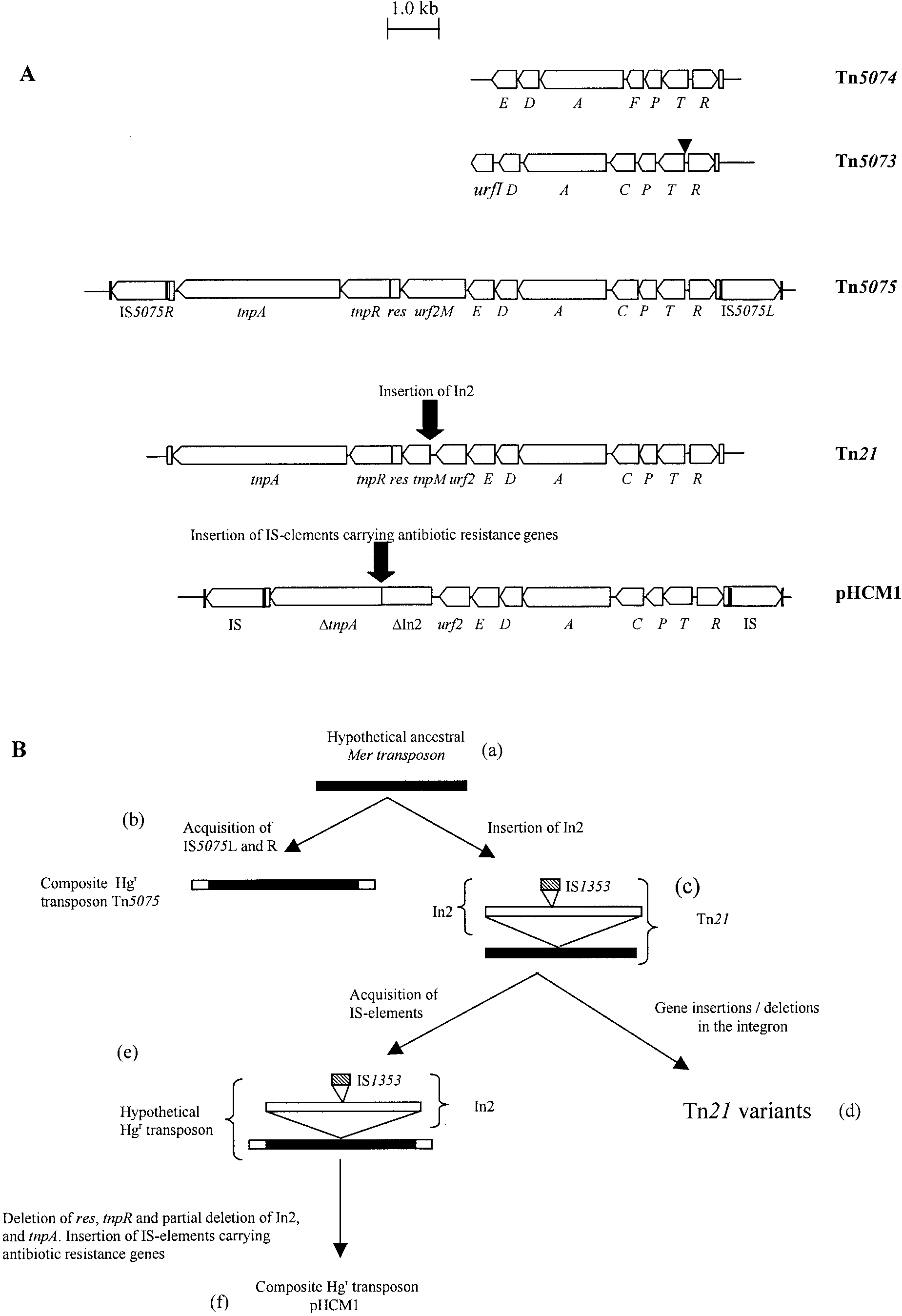 TABLE 2. Comparison of DNA sequence identities between Tn5073, Tn5074, Tn5075, and
published DNA sequences from other transposons
% Identity to mer transposon (genes)a
Ͼ99.9 Tn5036-like transposon pHCM1
Ͼ99.9 Tn21-like transposon from pHCM1 (merRTPCADE)
Ͼ99.8 Tn21 (res tnpA tnpR)
Ͼ97.1 hypothetical urf2m from Tn21⌬
Ͼ99.6 Tn21-like transposon from pHCM1 (IS elements)
Ͼ91.0 IS4321L, IS4321R, from R751
cell. We found no transposition of Tn5075 in our assays, which
are as follows: Tn5073 (strain M426), AF461013; Tn5074 (strain
could detect frequencies of Ͼ10Ϫ7.
TABLE 2. Comparison of DNA sequence identities between Tn5073, Tn5074, Tn5075, and
published DNA sequences from other transposons
% Identity to mer transposon (genes)a
Ͼ99.9 Tn5036-like transposon pHCM1
Ͼ99.9 Tn21-like transposon from pHCM1 (merRTPCADE)
Ͼ99.8 Tn21 (res tnpA tnpR)
Ͼ97.1 hypothetical urf2m from Tn21⌬
Ͼ99.6 Tn21-like transposon from pHCM1 (IS elements)
Ͼ91.0 IS4321L, IS4321R, from R751
cell. We found no transposition of Tn5075 in our assays, which
are as follows: Tn5073 (strain M426), AF461013; Tn5074 (strain
could detect frequencies of Ͼ10Ϫ7.