Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Biomedsearch.com
Nucleic Acids Research, 1997, Vol. 25, No. 61219–1224 Interaction of tetracycline with RNA: photoincorporation into ribosomal RNA of Escherichia coli Rudolf Oehler, Norbert Polacek, Guenter Steiner1 and Andrea Barta*
Institute of Biochemistry, University of Vienna, Vienna Biocenter, Dr Bohrgasse 9/3, A-1030 Vienna, Austria and1Division of Rheumatology, Department of Internal Medicine III, University of Vienna, Austria
Received November 18, 1996; Revised and Accepted February 3, 1997
ABSTRACT
the main target in experiments using [3H]tetracycline as aphotoaffinity reagent (5). Photolysis of [3H]tetracycline in the presence of
In previous experiments, we have used a photoreactive
Escherichia coli ribosomes results in an approximately
benzophenone derivative of tRNA [3-(4′-benzoylphenyl)propionyl-
1:1 ratio of labelling ribosomal proteins and RNAs. In
phenylalanine transfer RNA (BP-Phe-tRNA)] to characterize the
this work we characterize crosslinks to both 16S and
peptidyltransferase region on the 50S subunit (8,9). The photoreac-
23S RNAs. Previously, the main target of photoincor-
tion with the 23S RNA was completely inhibited by tetracycline,
poration of [3H]tetracycline into ribosomal proteins
and tetracycline itself crosslinked efficiently to the loop V region
was shown to be S7, which is also part of the one
of 23S RNA (9,10). This was somewhat surprising since the data
strong binding site of tetracycline on the 30S subunit.
from several investigations suggested that tetracycline might
The crosslinks on 23S RNA map exclusively to the
incorporate mainly into ribosomal proteins (5,11,12). We there-
central loop of domain V (G2505, G2576 and G2608)
fore undertook a thorough analysis of the photoincorporation of
which is part of the peptidyl transferase region.
tetracycline into ribosomal RNAs under conditions optimized to
However, experiments performed with chimeric ribo-
avoid non-specific binding and labelling due to tetracycline
somal subunits demonstrate that peptidyltransferase
photoproducts. We show that tetracycline can be crosslinked to
activity is not affected by tetracycline crosslinked
16S RNA as well as to 23S RNA but not to 5S RNA. Activity data
solely to the 50S subunits. Three different positions
from the crosslinked subunits show that the inhibitory effect
are labelled on the 16S RNA, G693, G1300 and G1338.
results solely from the interaction of tetracycline with the small
The positions of these crosslinked nucleotides corre-
subunit. This suggests that tetracycline crosslinks to 16S RNA
late well with footprints on the 16S RNA produced
from the strong binding site and that it might act via interaction
either by tRNA or the protein S7. This suggests that the nucleotides are labelled by tetracycline bound to the strong binding site on the 30S subunit. In addition, our MATERIALS AND METHODS results demonstrate that the well known inhibition of tRNA binding to the A-site is solely due to tetracycline Materials crosslinked to 30S subunits and furthermore suggest
Tetracycline hydrocloride was purchased from Sigma, highly
that interactions of the antibiotic with 16S RNA might
purified tetracycline was a present from Dr George Ellestad
be involved in its mode of action.
(Wyeth-Ayerst, Pearl River, NY). [3H]Tetracycline was purchasedfrom New England Nuclear (0.5 mCi/µmol). All tetracycline
INTRODUCTION
solutions were stored frozen in the dark and replaced frequentlybecause the drug undergoes both thermal and photochemical
The antibiotic tetracycline inhibits binding of tRNA to ribosomes
degradation. 70S Ribosomes were prepared from Escherichia
(1). Specifically, it mainly influences binding to the A-site
coli MRE600 as described (13). 30S and 50S ribosomal subunits
although some effects on the binding constant of Ac-Phe-tRNA
to the P-site have also been observed (2,3). Tetracycline binds toa single strong binding site on the 30S ribosomal subunit as well
Photocrosslink experiments
as to a number of weaker sites on both, the 30S and 50S subunits(2,4–6). The precise mechanism of tetracycline inhibition is not
Photolysis experiments were performed using a short arc mercury
known, but it is generally assumed that inhibition is caused by
lamp (HBO 500 W/2 from OSRAM) having an output concentrated
binding of tetracycline to the strong binding site on the 30S
at 366 nm. Samples were irradiated in vertical tubes at a distance
subunit (2,5,6). In a series of experiments where single proteins
of ∼200 mm from the lamp in the outer focal point (average
were omitted from the 30S subunit it has been established that the
luminance 3000 cd/cm2). Filters were chosen in such a way that
high affinity site is dependent on the presence of 16S RNA and
any light below 300 nm was completely eliminated. All photolyses
the proteins S3, S7, S8, S14 and S19 (7). Of these proteins, S7 was
were performed in standard TMK buffer (20 mM Tris–HCl, pH
*To whom correspondence should be addressed. Tel: +43 1 79 515 3520; Fax: +43 1 79 515 3114; Email: andrea@bch.univie.ac.at
1220 Nucleic Acids Research, 1997, Vol. 25, No. 6
7.4, 100 mM KCl, 6 mM MgCl2, 0.4 mM EDTA and 2 mM DTE)
Determination of Ac-[3H]Phe-tRNA binding to the
at 0_C. For the identification of the labelled nucleotides highly
ribosomal A- and P-site
purified tetracycline was used for crosslinking.
For P-site binding, 0.2 pmol/µl 70S ribosomes were incubated inT20K100M6 buffer (20 mM Tris–HCl pH 7.4, 100 mM KCl, 6 mM
Distribution of [3H]tetracycline photocrosslinked to
MgCl2, 0.4 mM EDTA and 2 mM DTE) for 10 min at room
ribosomal proteins and ribosomal RNA
temperature in the presence of 0.2 pmol/µl Ac-[3H]Phe-tRNAand 0.1 µg/µl poly(U). For A-site binding 0.2 pmol/µl 70S
After photolysis, ribosome samples were separated into two equal
ribosomes were pre-incubated in T20K100M12 buffer (20 mM
parts. In one of them ribosomal RNA was degraded by RNase T1.
Tris–HCl pH 7.4, 100 mM KCl, 12 mM MgCl2, 0.4 mM EDTA
The ribosomal proteins were precipitated by addition of 0.1 vol
and 2 mM DTE) for 3 min at 37_C in the presence of 0.2 pmol/µl
100 g/l BSA and 1 vol 10% TCA, redissolved in 10 M urea and
uncharged tRNAPhe and 0.1 µg/µl poly(U). Then 0.2 pmol/µl
TCA precipitated again. The precipitate was filtered through a
Ac-[3H]Phe-tRNA was added and the sample was incubated for
GF/C (Millipore) filter and washed several times with diethyl-
10 min at room temperature. To remove unbound Ac-[3H]Phe-
ether/ethanol (10:1) to remove unbound [3H]tetracycline. For
tRNA the samples were filtered through a nitrocellulose filter
determination of the amount of [3H]tetracycline photoincorporated
(NC-Filter, 45µm, Milipore, Molsheim, France). The filter was
into ribosomal RNA, the RNA was isolated by phenol/chloro-
washed several times with T20K100M6 or T20K100M12, respectively.
form extraction and precipitated. The pellet was dissolved in
The radioactivity of the filter corresponded to the bound
water and the radioactivity was measured. Virtually no background
of [3H]tetracycline was detectable in non-irradiated control samples. Determination of peptidyltransferase activity Reverse transcriptase analysis
The puromycin reaction, the formation of Ac-[3H]Phe-puromycin
RNA isolated from the ribosomes as described above was used
from Ac-[3H]Phe-tRNA and puromycin, was used to measure
for reverse transcriptase analysis according to (15). Primers used
peptidyltransferase activity. Ac-[3H]Phe-tRNA was bound to the
to investigate crosslinks on 16S and on 23S rRNA were the same
P-site as described above and incubated with 1 mM puromycin
for 10 min at room temperature. After addition of 1 vol 0.3 MNa-acetate (pH 5.5) in saturated MgSO4 the Ac-[3H]Phe-puromycin was extracted with ethyl acetate and the radioactivity
Synthesis of Ac-[3H]Phe-tRNA
measured in a scintillation counter.
tRNA was charged and acetylated as described (17). Ac-[3H]Phe-
tRNA was purified by reversed phase high performance liquidchromatography on nucleosil 300-5-C4-column (4 × 250 mm). Photoincorporation of [3H]tetracycline into E.coli ribosomes
Up to 30 nmol was typically applied to the column. The elutingsolvent had constant 400 mM NaCl, 10 mM Mg(CH
In the present work we reinvestigated the interaction of tetracy-
cline with ribosomal RNA. First, we determined the distribution
4-acetate, pH 5.0. The gradient steps had the following
percentages of methanol: 0%, 5 min; 0–9% in 5 min; 9–25% in
of tetracycline photocrosslinked to ribosomal RNA and proteins
50 min; 25%, 5 min. The different tRNA species were separated
using [3H]tetracycline as photoaffinity label. 70S ribosomes were
in the linear gradient from 9 to 25% of methanol. The fractions
irradiated in the presence of 100 µM [3H]tetracycline for different
containing Ac-[3H]Phe-tRNA were collected, desalted using an
periods of time. As shown in Figure 1, the amount of tetracycline
Econopac P6 desalting column (BioRad) and dried in a speedvac.
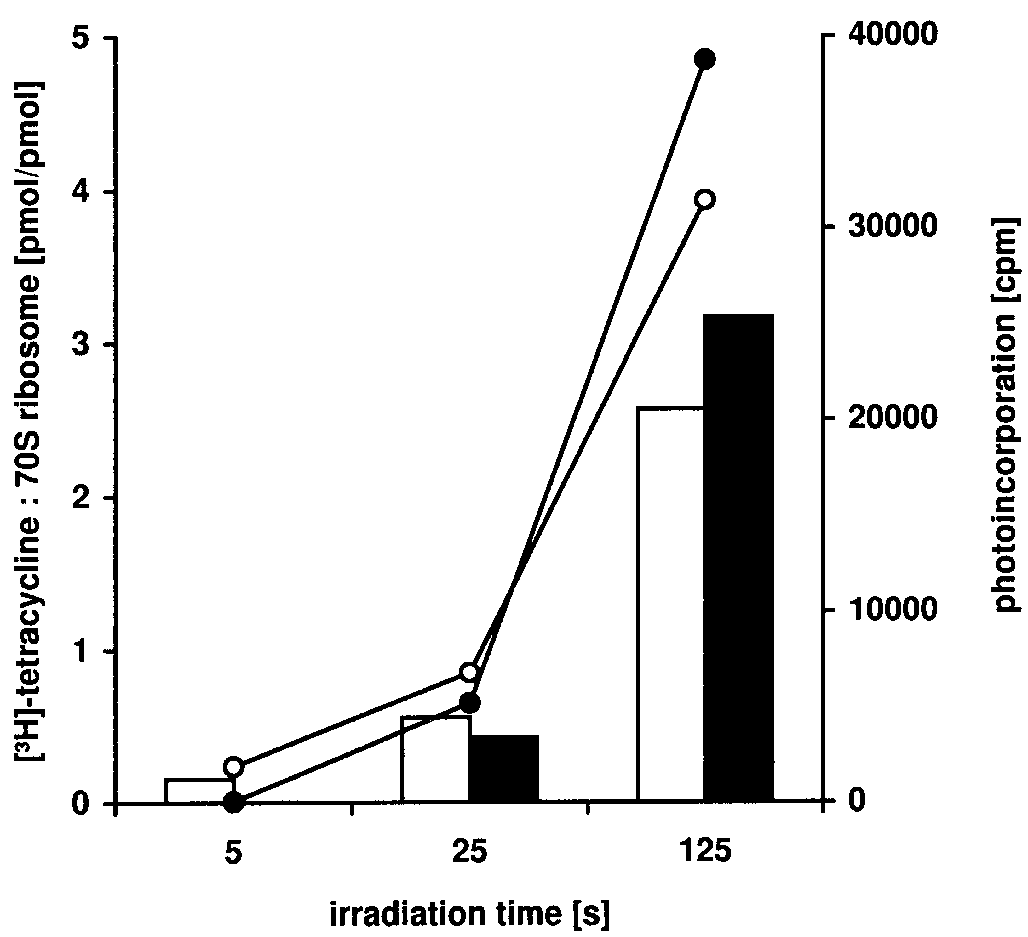
photocrosslinked to RNA and proteins increased with increasingirradiation time. Tetracycline was photocrosslinked to both, RNAand proteins, to about the same extent (Fig. 1). This is in contrast
Preparation of chimeric ribosomes
to the results of Goldman and co-workers (5,11) who only foundup to 10% of the tetracycline label incorporated into rRNA (see
Isolated 30S and 50S subunits were irradiated in the presence of
Discussion). Using 25 s irradiation time, tetracycline was already
tetracycline as described above. To remove unbound tetracycline,
incorporated into 70S ribosomes at an approximate ratio of 1:1.
the subunits were pelleted twice (first 8 h at 31 000 r.p.m. for 30S
Therefore, and to avoid secondary reactions of tetracycline
or 5 h at 28 000 r.p.m. for 50S, then 18 h at 21 000 r.p.m. for 30S
photoproducts we used an irradiation time of just 30 s in all
or 18 h at 15 000 r.p.m. for 50S) in a Beckman ultracentrifuge
using the SW50.1 rotor. The pellet was dissolved in T20M20N400buffer (20 mM Tris–HCl pH 7.5, 20 mM MgCl2, 400 mM NH4Cl,
Localization of tetracycline–rRNA photocrosslinks
4 mM β-mercaptoethanol). After addition of an equimolaramount of the complementary untreated ribosomal subunit the
The analysis of the ribosomal proteins photolabelled by [3H]tetra-
samples were incubated for 10 min at 37_C. The samples were
cycline confirmed the results of Goldman et al. (5), as we could
then centrifuged in a 10–30% sucrose gradient in T20M10N100
also identify protein S7 as the major labelled protein (data not
buffer (20 mM Tris–HCl pH 7.5, 10 mM MgCl2, 100 mM NH4Cl,
shown). We therefore concentrated on the analysis of the sites of
6 mM β-mercaptoethanol, 0.5 mM EDTA) for 18 h at
tetracycline–rRNA interactions using the primer extension
18 000 r.p.m. (Beckman SW28 rotor). The fractions containing
method. The primers chosen were spaced every ∼200 nucleotides
70S chimeric ribosomes were collected and centrifuged for 24 h
on the 16S, 23S and 5S RNA, so we were able to scan the entire
at 24 000 r.p.m. (Beckman 45Ti rotor). The pellet was resuspended
RNAs except the 3′-ends. The RNAs used for the templates were
in T20M10N100 (containing 50% glycerol) and stored at –70_C.
from 70S ribosomes irradiated in the presence of different
1221 1221 Figure 1. Photoincorporation of [3H]tetracycline into E.coli ribosome. 70S Ribosomes (36 pmol) were irradiated in presence of 100 µM [3H]tetracycline in 100 µl TMK buffer for the indicated period of time. Lines indicate radioactivity found upon irradiation in ribosomal RNA (filled circles) and in ribosomal proteins (open circles). The boxes indicate pmoles of [#H]tetracy- cline photoincorporated per pmole ribosomal RNA (black boxes) and ribosomal proteins (open boxes). Values are given as mean of four parallel experiments.
amounts of tetracycline (Fig. 2). RNAs from non-irradiated 70Sribosomes and from ribosomes irradiated in the absence oftetracycline were used as controls for random stops on the RNAtemplate and possible UV-induced internal RNA–RNA cross-links. When a stop was observed, the crosslinked nucleotide wastaken to be the following nucleotide in the rRNA template (i.e. thepreceding one in the rRNA sequence). The numbers of photoaffinitylabelled nucleotides increased with rising concentrations oftetracycline. The half maximal inhibition of Ac-Phe-tRNA
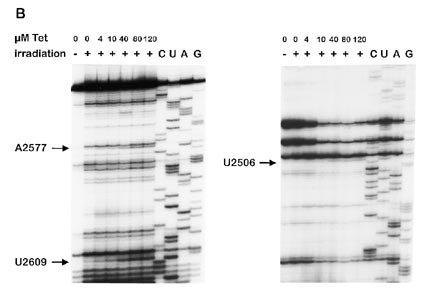
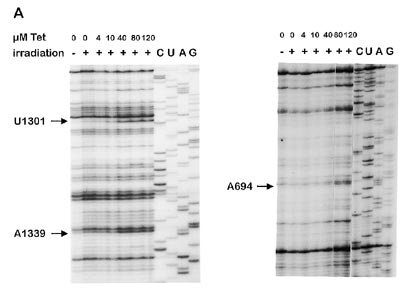
Figure 2. Primer extension experiments showing reverse transcriptase
binding to the ribosome by tetracycline was reported to be 40 (18)
elongation stops caused by photocrosslinks of tetracycline to ribosomal RNAs.
or 4 µM (3), respectively, whereas under our incubation
A suitable set of oligonucleotide primers were used for analysing 16S RNA (A) and 23S RNA (B). Only those crosslinks are indicated which are already
conditions we observed a value of ∼10 µM. Therefore, only those
nucleotides were considered to correlate well with the inhibitoryaction of tetracycline which were labelled in the presence of40 µM or lower concentrations of the antibiotic. Under theseconditions, three sites on the 16S RNA (Fig. 2A; G693, G1300
subunits in the presence of 40 µM tetracycline produced the same
and G1338) and three sites on the 23S RNA (Fig. 2B; G2505,
RNA labelling pattern as irradiation of 70S ribosomes (data not
G2576 and G2608) were photoaffinity labelled by tetracycline.
shown). This indicated that the 50S crosslinks originated from a
No incorporation of tetracycline into 5S RNA could be detected
tetracycline binding site on the 50S subunit.
(data not shown). Several additional labelled nucleotides on 16Sand 23S RNA could be identified when 80 or 120 µM tetracycline
Effect of photoincorporated tetracycline on ribosomal
were used and some of them are discussed later. function
The positions of the labelled nucleotides are shown in Figure
3A in a two dimensional model of the 16S RNA. In addition, we
Next we investigated the effect of tetracycline photocrosslinks on
have indicated nucleotides which produce footprints with tRNA
ribosomal function using chimeric ribosomes. To distinguish the
(19), protein S7 (20) and tetracycline (21). As can be seen in
effects of tetracycline crosslinked to the 30S subunit from those
Figure 3A, the labelled nucleotides are close to nucleotides either
of tetracycline crosslinked to the 50S subunit we performed the
involved in binding of tRNA or protein S7.
following experiments. Isolated 30S or 50S ribosomal subunits
On the 23S RNA the labelled nucleotides were located
were irradiated in the presence of tetracycline. Then we
exclusively in the central loop region of domain V (Fig. 3B). This
immediately removed the unbound tetracycline by centrifugation.
loop has been identified as an essential part of the peptidyltrans-
The ribosomal subunits were reconstituted to 70S particles with
ferase region on the 50S subunit [for reviews see 10, 22)]. As the
the complementary untreated subunit. The chimeric ribosomes
main binding site of tetracycline had been located on the 30S
were then isolated by density gradient centrifugation and the
subunit we wondered if the photocrosslinks to the 50S subunit
effects of the photomodified subunits on binding of peptidyl-tRNA
were derived from tetracycline bound to the main binding site on
to the ribosome and on peptidyltranferase activity were investigated.
the 30S subunit which might be located at the interface between
As shown in Figure 4, the photocrosslinked 50S ribosomal
the 30S and 50S subunit. However, irradiation of isolated 50S
subunit had no effect on binding of Ac-[3H]Phe-tRNA either to
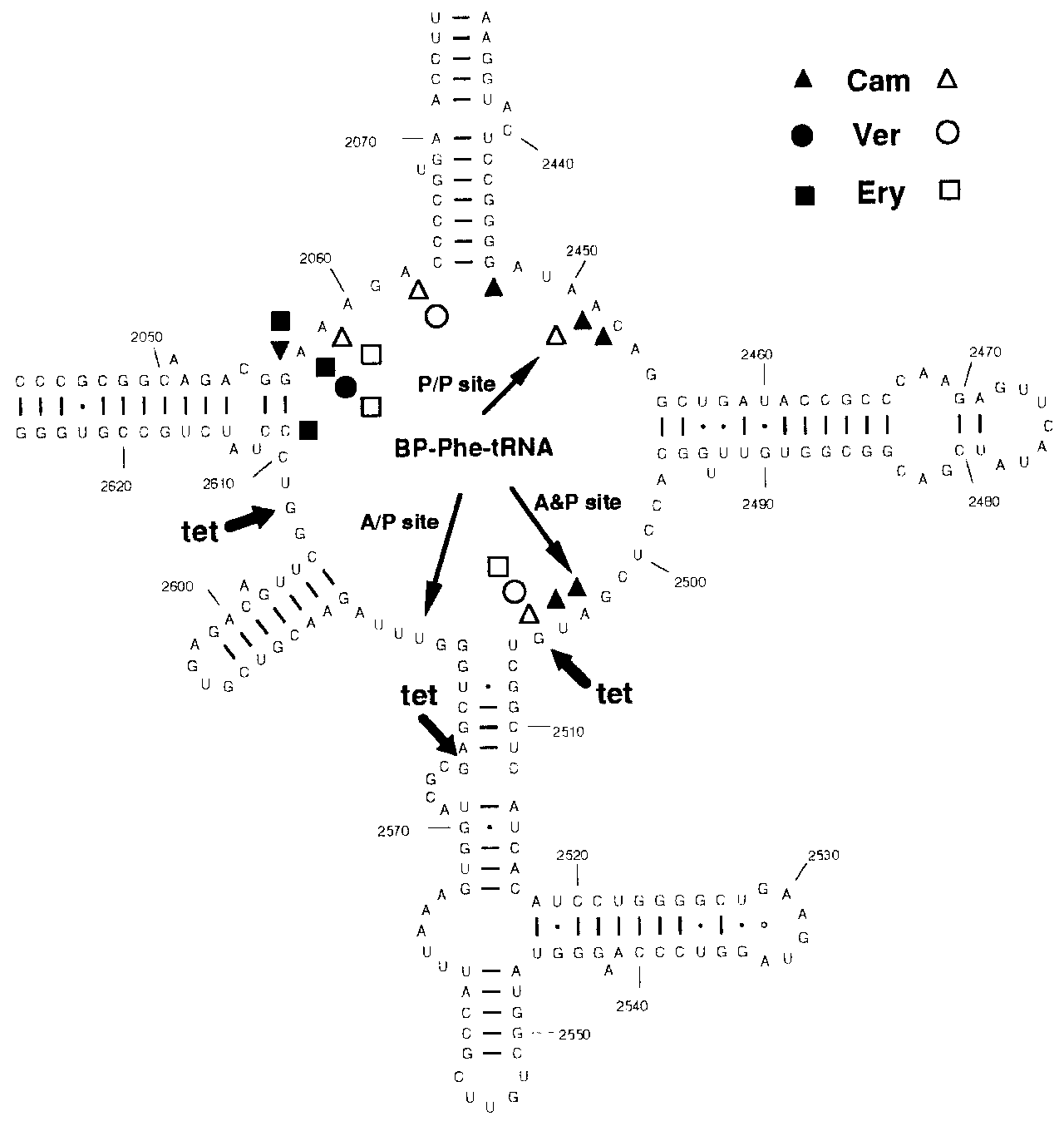
1222 Nucleic Acids Research, 1997, Vol. 25, No. 6 Figure 3. (A) Secondary structure diagram of 16S rRNA. Positions of nucleotides photoaffinity labelled by 40 µM tetracycline are indicated by thick arrows. On the 16S rRNA diagram bases protected from chemical modification by binding of protein S7 (filled circles) (30), of tRNA (triangles) (19) and of tetracycline (open circle) (21) are indicated in addition to nucleotides which have enhanced reactivities towards chemical probing when tetracycline is bound to the ribosome (thin arrows) (21). (B) Secondary structure of the central loop region of domain V of 23S rRNA. Positions of nucleotides photoaffinity labelled by 40 µM tetracycline are indicated by thick arrows. The nucleotides specifically labelled by A- and P-site bound BP-Phe-tRNA are indicated by the thin arrows (9). Cam, chloramphenicol; Ery, erythromycin; Ver, vernamycin. Filled symbols indicate nucleotides whose mutation confers resistance to the respective antibiotic (31–34) open symbols designate antibiotics whose binding to the ribosome causes an alteration of reactivity of the respective nucleotide toward chemical modification (35).
the ribosomal A- or to the P-site when compared with untreated
30 W and irradiation times between 60 and 90 min (108–162 kJ)
(11). These workers also showed that upon irradiation tetracycline
The Ac-[3H]Phe-puromycin formation, which measures peptidyl-
photoproducts were generated which could further react with the
tranferase activity, was also unaffected by tetracycline photo-
ribosome (5). Therefore, we took care to avoid long irradiation
crosslinked to the 50S subunit. In contrast, chimeric ribosomes
times. Furthermore, as our results did not change upon addition
with tetracycline photocrosslinked to the 30S subunit had
of β-mercaptoethanol which has been used to avoid light
diminished ability of binding Ac-[3H]Phe-tRNA to the ribosomal
independent incorporation of tetracycline photoproducts and as
A-site compared with untreated 70S ribosomes, whereas the
our protein labelling pattern conforms to the one published
P-site binding and the Ac-[3H]Phe-puromycin formation remained
previously (5), we are confident that the crosslinks observed
unaffected. These results correlate well with the published data on
the inhibition of A-site binding of tRNA by tetracycline (2,3).
The experiments were performed with increasing concentrations
of tetracycline; however, only those crosslinks have been
DISCUSSION
described which appear v40 µM tetracycline. The number of
The experiments described in this paper show that tetracycline
crosslink sites increased with higher concentrations (e.g. on 23S
can be photocrosslinked not only to ribosomal proteins, but also
RNA three more at 80 µM, and additional seven at 120 µM
to rRNA. We found an approximately 1:1 incorporation of
tetracycline), in accordance with a large number of low affinity
radioactivity in ribosomal proteins and RNA, respectively.
binding sites for tetracycline observed on both the 30S and 50S
[3H]Tetracycline has been previously used in extensive studies to
characterize ribosomal binding sites for this antibiotic (5,11,12).
Previously, the strong binding site of tetracycline was localized
In these experiments up to 90% of the radioactivity was found to
to the 30S ribosomal subunit (4–6). There are several results
be incorporated in ribosomal proteins with S7 being the main
supporting the idea that the inhibitory effect of tetracycline on
protein labelled (5). The difference in the distribution of the label
protein synthesis, i.e. the blocking of aminoacyl-tRNA binding to
may result from the different irradiation conditions used. We used
the A-site, is a direct consequence of its binding to the strong
∼500 W for only 30 s (15 kJ) whereas Cooperman’s group used
binding site (4–6,23). In addition, it has been shown that 16S
1223 1223
located at the interface between the two ribosomal subunits. Given the location of the crosslinks, we were surprised not to findany effect on peptidyltransferase activity. In our previousexperiments using BP-Phe-tRNA as affinity label for thepeptidyltransferase region, we observed an inhibition of theBP-Phe-tRNA crosslinks, but not of its binding, by severalantibiotics including tetracycline and chloramphenicol (9). Oneexplanation of this observation would be that tetracycline mightbind to the peptidyltransferase region and distort the peptidyl endof tRNA thus inhibiting the BP-Phe-tRNA crosslink to the 23SRNA. Binding of tetracycline to this region is similar but notidentical to chloramphenicol (an inhibitor of peptidyltransferaseactivity) as peptidyltransferase activity in vitro is not inhibited. Although, an effect on ribosomal function in vivo cannot beexcluded, this effect would be overruled by the effect oftetracycline on binding of tRNA to the A-site.
The labelled nucleotides of the 16S rRNA are shown in Figure
3A together with footprints created by binding of tRNA, proteinS7, and tetracycline. The distribution of all these nucleotidesdemonstrates that the photoaffinity labelled nucleotides (G693,G1300 and G1338) on the 16S RNA are close to footprint sitesof tRNA and protein S7. Furthermore, G890 which is labelled by120 µM tetracycline (data not shown) is adjacent to nucleotideA892 which was protected upon binding of 100 µM tetracycline
Figure 4. Effects of photocrosslinked tetracycline on ribosomal function.
to chemical probing (Figure 3A; 21). It is interesting to note that
Tetracycline was photocrosslinked to isolated 30S and 50S ribosomal subunits.
several of the other nucleotides labelled additionally at higher
After removing the unbound tetracycline the subunits were reconstituted to 70Sribosomes by addition of untreated 50S subunits and 30S subunits, respectively.
concentrations of tetracycline are also located near tRNA
The diagrams compare these chimeric ribosomes with ribosomes, which were
footprint sites (data not shown). As 16S RNA and the protein S7
reconstituted from a subunit irradiated in the absence of tetracycline and an
are essential components of the strong binding site of tetracycline
untreated subunit. Ac-[3H]Phe-tRNA binding to the A-site and P-site and
on the 30S subunit, it is likely that the labelling of the nucleotides
Ac-[3H]Phe-puromycin formation are shown.
on the 16S RNA occurs from the main binding site. It is not
RNA together with the proteins S3, S7, S8, S14 and S19 are
known how tetracycline inhibits binding of tRNA to the A-site
essential for providing the binding domain for tetracycline on the
but our data reveal that the close proximity of tetracycline to the
30S subunit and that within this domain S7 is the major labelled
16S RNA may be one of the determinants of its mode of action.
protein (5,7). Although we observed an RNA to protein labelling
Thus tetracycline might act by interfering with the tRNA/16S
ratio different from that seen by other investigators, we too found
RNA interaction directly or via a structural distortion of the 16S
the same protein labelling pattern with protein S7 being the main
RNA brought about by binding of tetracycline to its high affinity
protein labelled (data not shown). Tetracycline has been found to
binding site with its main contacts to S7 and 16S RNA. Distortion
bind either to proteins, such as the Tet repressor (24), or to RNA
of 16S RNA was also implicated by the enhancement of the
as was demonstrated for group I and group II introns (25). As no
reactivities of U1052 and C1054 to chemical probing upon
binding to ribosomal proteins free in solution occurs (26) there is
binding of tetracycline (21). A thorough analysis of the action of
the possibility of a mixed RNA–protein binding site on the
streptomycin and neomycin has recently led to a model putting
ribosome. In order to characterize this site, the crosslinked
forward the idea that these antibiotics act by distortion of 16S
nucleotides on the ribosomal RNAs were analysed.
RNA structures (29). These antibiotics which give different
Both 16S and 23S rRNAs were photoaffinity labelled by
footprints to 16S RNA than tetracycline have an influence on the
tetracycline. Interestingly, 23S RNA was labelled exclusively in
decoding fidelity. Tetracycline, on the other hand, might have an
the central loop of domain V, the peptidyltransferase centre (Fig.
effect on the high affinity binding of tRNA after the decoding
3B), but this labelling did not affect tRNA binding and peptidyl
transfer. These crosslinks originate from a binding site on the 50Ssubunit as the same crosslinks were observed when merely 50Ssubunits were used. At 80 µM tetracycline two additional
ACKNOWLEDGEMENTS
crosslinks were found in this region; some of them havepreviously been identified in crosslink experiments where100 µM tetracycline has been used (9). Therefore, on the 50S
We would like to thank I. Halama and S. Dorner for help during
subunit tetracycline seems to bind solely to the peptidyl
preparation of the manuscript, B. Weiser for the XRNA program,
transferase region at least at lower concentrations. It is interesting
R. Schroeder, K. Nierhaus and Z. Rattler for invaluable discussions
and probably not by chance that tetracycline crosslinks mainly to
and infinite patience. This work was supported by a grant
the peptidyl transferase region on 23S RNA where various other
(P09454-MIB) from the ‘Österreichischer Fonds zur Förderung
antibiotics like chloramphenicol and erythromycin bind (Fig.
der wissenschaftlichen Forschung’ and from the ‘Anton Dreher
3B). This region is mainly composed of RNA (8,27,28) and
Gedächtnisschenkung für Medizinische Forschung’ to A.B. 1224 Nucleic Acids Research, 1997, Vol. 25, No. 6 REFERENCES
18 Suarez,G. and Nathans,D. (1965) Biochem. Biophys. Res. Commun., 18,
19 Moazed,D. and Noller,H.F. (1986) Cell, 47, 985–994.
1 Gottesmann,M.E. (1967) J. Biol. Chem., 242, 5564–5571.
20 Noller,H.F., Moazed,D., Stern,S., Powers,T., Allen,P.N., Robertson,J.M.,
2 Epe,B., Woolley,P. and Hornig,H. (1987) FEBS Lett., 213, 443–447.
Weiser,B. and Triman,K. (1990) In Hill,W.E., Dahlberg,A., Garrett,R.A.,
3 Geigenmuller,U. and Nierhaus,K.H. (1986) Eur. J. Biochem., 161,
Moore,P.B., Schlessinger,D. and Warner,J.R. (eds), The Ribosome:Structure, Function and Evolution. American Society for Microbiology,
4 Epe,B. and Woolley,P. (1984) EMBO J., 3, 121–126.
5 Goldman,R.A., Hasan,T., Hall,C.C., Strycharz,W.A. and Cooperman,B.S.
21 Moazed,D. and Noller,H.F. (1987) Nature, 327, 389–394.
(1983) Biochemistry, 22, 359–368.
22 Rodriguez Fonseca,C., Amils,R. and Garrett,R.A. (1995) J. Mol. Biol.,
6 Tritton,T.R. (1977) Biochemistry, 16, 4133–4138. 247, 224–235.
7 Buck,M.A. and Cooperman,B.S. (1990) Biochemistry, 29, 5374–5379.
23 Streltsov,S.A., Kukhanova,M.K., Krayevsky,A.A., Beljavskaja,I.V.,
8 Barta,A., Steiner,G., Brosius,J., Noller,H.F. and Kuechler,E. (1984) Proc.
Victorova,L.S., Gursky,G.V., Treboganov,A.D. and Gottikh,B.P. (1974)
Natl Acad. Sci. USA, 81, 3607–3611. Mol. Biol. Rep., 1, 391–396.
9 Steiner,G., Kuechler,E. and Barta,A. (1988) EMBO J., 7, 3949–3955.
24 Hillen,W. and Berens,C. (1994) Annu. Rev. Microbiol., 48, 345–369.
10 Barta,A., Kuechler,E. and Steiner,G. (1990) In Hill,W.E., Dahlberg,A.,
25 Liu,Y., Tidwell,R.R. and Leibowitz,M. J. (1994) J. Euk. Microbiol., 41,
Garrett,R.A., Moore,P.B., Schlessinger,D. and Warner,J.R. (eds), TheRibosome: Structure, Function and Evolution. ASM, Washington, DC, pp.
26 Buck,M.A., Olah,T.V., Perrault,A.R. and Cooperman,B.S. (1991)
Biochimie, 73, 769–775.
11 Goldman,R.A., Cooperman,B.S., Strycharz,W.A., Williams,B.A. and
27 Barta,A. and Kuechler,E. (1983) FEBS Lett, 163, 319–323.
Tritton,T.R. (1980) FEBS Lett., 118, 113–118.
28 Noller,H.F. (1993) FASEB J., 7, 87–89.
12 Reboud,A.M., Dubost,S. and Reboud,J.P. (1982) Eur. J. Biochem., 124,
29 Karimi,R. and Ehrenberg,M. (1996) EMBO J., 15, 1149–1154.
30 Stern,S., Powers,T., Changchien,L.M. and Noller,H.F. (1989) Science, 244,
13 Moazed,D. and Noller,H.F. (1989) Cell, 57, 585–597.
14 Rheinberger,H.J., Geigenmuller,U., Wedde,M. and Nierhaus,K.H. (1988)
31 Douthwaite,S., Prince,J.B. and Noller,H.F. (1985) Proc. Natl. Acad. Sci.Methods Enzymol., 164, 658–670. USA, 82, 8330–8334.
15 Kuechler,E., Steiner,G. and Barta,A. (1988) Methods Enzymol, 164,
32 Ettayebi,M., Prasad,S.M. and Morgan,E.A. (1985) J. Bacteriol., 162,
16 Stern,S., Moazed,D. and Noller,H.F. (1988) Methods Enzymol., 164,
33 Noller,H.F. (1984) Annu. Rev. Biochem., 53, 119–162.
34 Sor,F. and Fukuhara,H. (1984) Nucleic Acids Res., 12, 8313–8318.
17 Kuechler,E. and Barta,A. (1977) Methods Enzymol., 46, 676–683.
35 Moazed,D. and Noller,H.F. (1987) Biochimie, 69, 879–884.
Dr Charlie Hamilton MBChB DCH nMRCGP (pending) PERSONAL DETAILS Name: Date of birth: Nationality: Contact Address: Telephone No.: charliehamilton@doctors.org.uk; charliehamilton@nhs.net 6102719 MDU No.: Current position: GP Registrar to Grosvenor Road Surgery, Paignton _____________________________________________________________________________
Atomic Lithium Beam Spectroscopy for N and T in Reactive Plasmas Institut für Experimentalphysik II, Ruhr-Universität Bochum, D-44780 Bochum, Germany 1. Introduction In a reactive plasma the species composition and the flows of particles to a substrate depend upon the electron energy distribution and its spatial variation and time dependence. The electron density has a large influence on
 1221
1221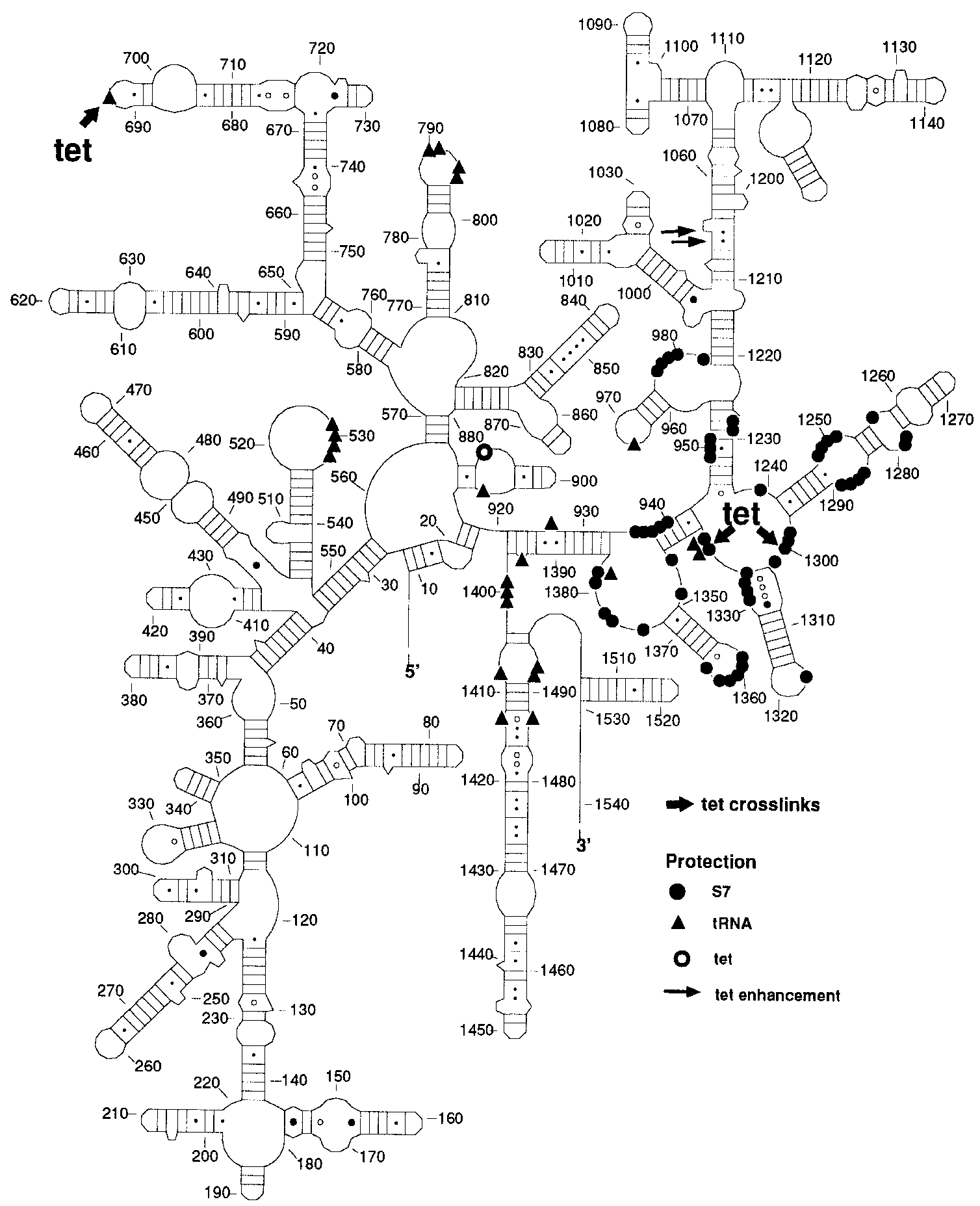
 1222 Nucleic Acids Research, 1997, Vol. 25, No. 6
1222 Nucleic Acids Research, 1997, Vol. 25, No. 6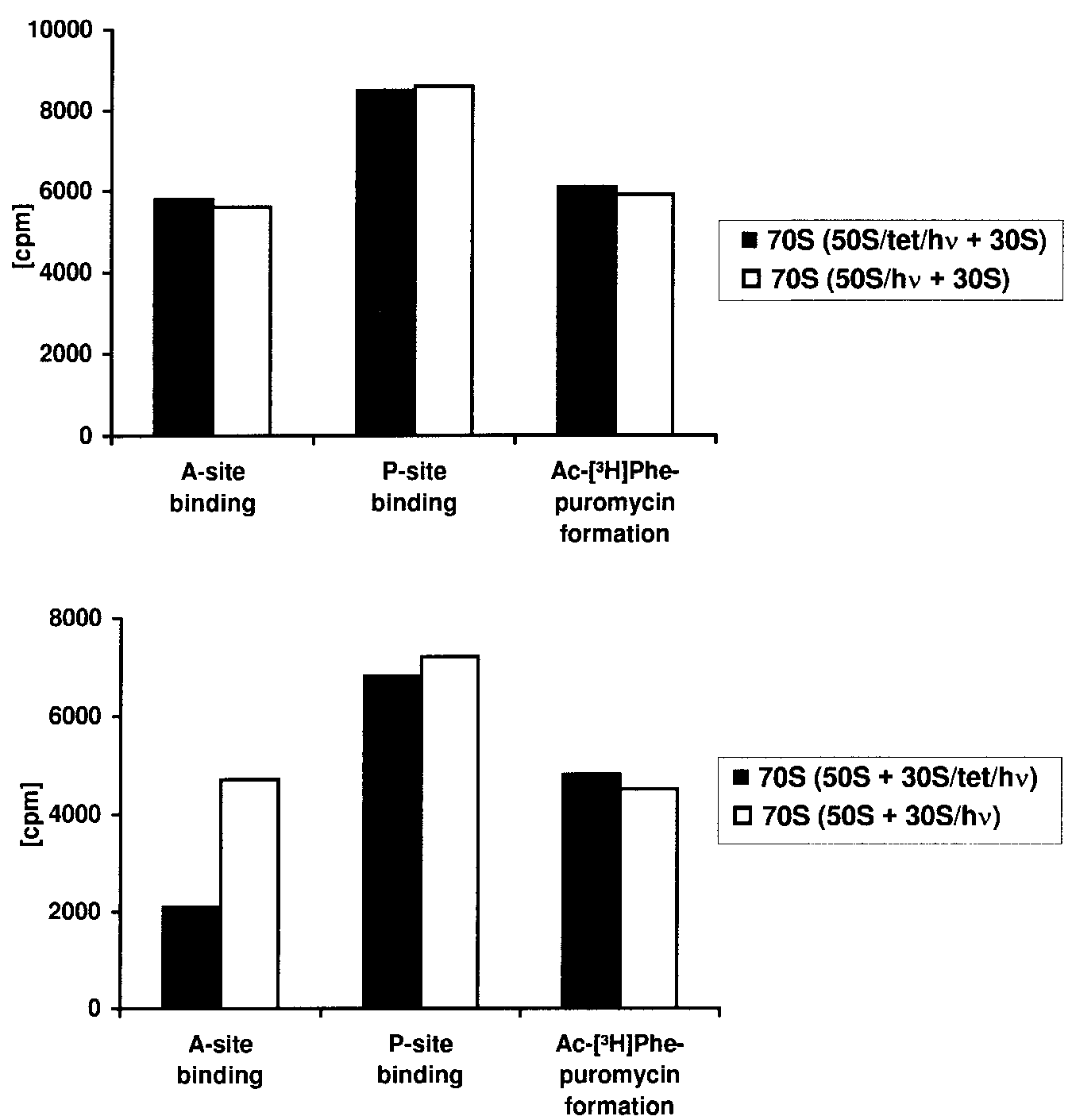 1223
1223