Le sildénafil présent dans Kamagra exerce une inhibition réversible de la PDE5, modulant la cascade GMPc et favorisant une vasodilatation localisée. L’absorption digestive varie selon la forme utilisée, comprimés classiques ou gels oraux. La distribution tissulaire est large et la liaison protéique élevée, avoisinant 96 %. La métabolisation hépatique génère un métabolite actif contribuant à l’effet pharmacologique global. La demi-vie reste courte, avec disparition plasmatique en quelques heures. Les interactions significatives concernent surtout les nitrés organiques et inhibiteurs puissants du CYP3A4. Dans les publications techniques, kamagra en ligne est souvent cité dans le cadre d’analyses comparatives portant sur les différences de formulations et de cinétique d’absorption.
Bcwgpsp.enfermedadesraras.es
Bone marrow transplantation restores immune system function and preventslymphoma in Atm-deficient mice
Jessamyn Bagley, Maria L. Cortes, Xandra O. Breakefield, and John Iacomini
Ataxia-telangiectasia (A-T) is a human jor causes of morbidity and mortality in regimen can be used to overcome the autosomal recessive disease caused by A-T patients. In mice, an introduced muta- immune deficiencies and prevent the mutations in the gene encoding ataxia- tion in Atm leads to a phenotype that malignancies observed in these mice. telangiectasia mutated (ATM). A-T is char- recapitulates many of the symptoms of Therefore, bone marrow transplantation acterized by progressive cerebellar de- A-T, including immune system abnormali- may prove to be of therapeutic benefit in generation, variable immunodeficiency, ties and susceptibility to malignancy. Here A-T patients. (Blood. 2004;104:572-578) and a high incidence of leukemia and we show that the replacement of the bone lymphoma. Recurrent sino-pulmonary in- marrow compartment in Atm knockout fections secondary to immunodeficiency mice (Atm؊/؊) using a clinically relevant, and hematopoietic malignancies are ma- nonmyeloablative host–conditioning 2004 by The American Society of Hematology Introduction
Ataxia telangiectasia (A-T) is a human autosomal recessive disease
CD4ϩCD8ϩ double-positive and CD4ϪCD8Ϫ double-negative thy-
that affects between 1 in 40 000 and 1 in 100 000 persons
mocytes is increased, whereas the frequency of CD4 and CD8
worldwide and is characterized by a wide variety of clinical
single-positive mature thymocytes is decreased when compared
manifestations.1,2 A-T is caused by mutations in a single gene,
with healthy mice,9-11 suggesting that Atm may be required for the
encoding ataxia-telangiectasia mutated (ATM). Symptoms of A-T
transition of immature CD4ϩ8ϩ double-positive thymocytes to the
include progressive cerebellar degeneration manifested mainly as
mature single-positive stage. It has been suggested that this
ataxia, oculocutaneous telangiectasias, recurrent pulmonary infec-
apparent block in T-cell development may also result in a marked
tions caused by immunodeficiency, lymphoreticular malignancies,
reduction in the number of mature CD4 and CD8 T cells in the
growth retardation, incomplete sexual maturation, and premature
periphery.10 In A-T patients, it has been reported that although total
aging of the skin and hair.3 The disease is progressive, and death
T-cell numbers in the blood are similar to those observed in healthy
generally occurs by the second or third decade of life. Hematologic
persons, the frequency of naive T cells is reduced, and the
malignancies, such as leukemia and lymphoma, can occur in as
frequency of memory marker–positive T cells is increased.12-14
many as 40% of patients4 and, together with bronchial infections,
A-T patients exhibit thymic hypoplasia, resulting in decreased
are the major causes of death in A-T patients. Defects in the
T-cell production and immunodeficiency, and hematologic malig-
immune system include decreased immunoglobulin A (IgA), IgE,
nancy. These abnormalities may result from defects intrinsic to
and IgG2 production, marked thymic hypoplasia, and defects in
hematopoietic stem cells (HSCs), or they may reflect developmen-
T-cell–mediated responses.3 Patients with A-T have extreme radia-
tal defects in the thymic microenvironment in which the progeny of
tion sensitivity and decreased tolerance to chemotherapeutic agents,
these cells mature. Defects in thymic function, such as those
preventing the use of standard therapies to treat malignancy.5-7
observed in DiGeorge syndrome, are known to result in immunode-
There is no cure for A-T; hence, treatments are directed toward
ficiency (for a review, see Buckley15). It has also been suggested
that fetal thymus transplantation may reverse the immunodefi-
AtmϪ/Ϫ mice, created by gene targeting, display many of the
ciency observed in A-T by overcoming thymic hypotrophy (for a
hallmarks of A-T seen in humans, including growth retardation,
review, see Saha and Chopra16). In addition, although thymic
infertility, defects in T-lymphocyte maturation, extreme sensitivity
development of T cells is impaired in A-T patients, the function of
to ␥-irradiation, and high incidence of hematologic malignancy.8-11
mature T cells has been reported to be normal,12,17 suggesting either
Mice in most Atm-deficient strains acquire malignant thymic
that a development-specific defect exists in T-cell progenitors or
lymphomas between 2 and 4 months of age and generally die
that the thymic microenvironment is unable to mediate efficient
before 30 weeks of age.9 AtmϪ/Ϫ mice also exhibit aberrant T-cell
T-cell maturation. We hypothesized that if there were intrinsic
development characterized by a decrease in absolute numbers of
defects in the HSCs of AtmϪ/Ϫ mice, replacing the hematopoietic
thymocytes. In the thymi of AtmϪ/Ϫ mice, the frequency of
compartment in these mice by bone marrow transplantation (BMT)
From the Transplantation Biology Research Center, Massachusetts General
Reprints:
Hospital and Harvard Medical School; and Molecular Neurogenetics Unit,
Massachusetts General Hospital, MGH-East, 149-5210 13th St, Boston, MA
Department of Neurology, Massachusetts General Hospital and Harvard
02129; e-mail: john.iacomini@tbrc.mgh.harvard.edu.
The publication costs of this article were defrayed in part by page charge
Submitted December 11, 2003; accepted March 2, 2004. Prepublished online as
payment. Therefore, and solely to indicate this fact, this article is hereby
Blood First Edition Paper, March 25, 2004; DOI 10.1182/blood-2003-12-4226.
marked ‘‘advertisement’’ in accordance with 18 U.S.C. section 1734.
Supported in part by a grant from the A-T Children’s Project and by NationalInstitutes of Health grants ROI AI43619-05 (J.I.) and T32 AI07529 (J.B.).
2004 by The American Society of Hematology
BLOOD, 15 JULY 2004 ⅐ VOLUME 104, NUMBER 2
REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
would overcome the observed hematologic abnormalities. Ourresults indicate that full donor-type hematopoiesis can be achieved
in AtmϪ/Ϫ mice using clinically relevant host conditioning, result-ing in the restoration of normal immune system function. In
Defects in lymphocyte development observed in Atm؊/؊ mice
addition, replacing the Atm-deficient hematopoietic compartment
are stem cell intrinsic
prevents the development of hematologic malignancies in Atm-
To determine whether defects in T-cell development observed in
deficient mice. Therefore, BMT may prove to be of significant
AtmϪ/Ϫ mice were caused by defects in the ability of the thymic
therapeutic benefit in A-T patients.
environment to support T-cell maturation, we monitored thedevelopment of AtmϪ/Ϫ mutant–derived T cells in wild-type micewith normal thymi. Wild-type C3H (H-2k) mice were lethallyirradiated and reconstituted with either 107 AtmϪ/Ϫ (H-2b) or
Materials and methods
wild-type littermate (Atmϩ/ϩ) control bone marrow cells. BothAtmϪ/Ϫ and Atmϩ/ϩ bone marrow cells efficiently engrafted in
lethally irradiated C3H recipients, resulting in more than 99%
AtmϪ/Ϫ knockout mice used as bone marrow donors for reconstitution of
donor-type cells in the blood at 6 weeks after BMT (Figure 1).
C3H recipients were a kind gift from Dr Fred Alt (Children’s Hospital,
Engraftment of donor bone marrow was stable, and multihemato-
Boston, MA). Mice were obtained as heterozygotes and were intercrossed
poietic lineage chimerism was maintained long term (Figure 1).
to obtain homozygous progeny that were genotyped by polymerase chain
Analysis of T-cell development in the thymi of recipients of
reaction (PCR) according to the protocol described.8 Heterozygous 129S6/
bone marrow transplants revealed defects in the ability of T-cell
SvEvTac-Atmtm1-Awb mice were purchased from the Jackson Laboratory
progenitors derived from AtmϪ/Ϫ HSCs to develop from the
(Bar Harbor, ME) and were used in all other experiments. Mice were
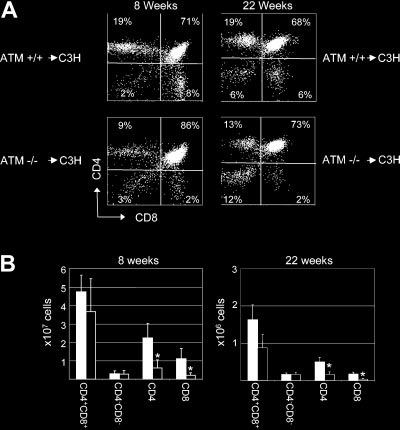
double-positive to the mature single-positive stage. Eight weeks
genotyped using PCR according to the manufacturer’s instructions (Jackson
after BMT, C3H mice reconstituted with AtmϪ/Ϫ bone marrow
Laboratory). C3H mice were obtained from a colony at Massachusetts
exhibited a block in T-cell development, resulting in an increase in
General Hospital. C3H mice are of the H-2k haplotype and are completely
the frequency of CD4ϩCD8ϩ double-positive thymocytes
major histocompatibility complex (MHC)–mismatched with 129S6/
(79% Ϯ 6%; n ϭ 8) when compared with the frequency observed
SvEvTac-Atmtm1-Awb mice, which are H-2b. B6.CH-2bm1 skin graft donors
in recipients of Atmϩ/ϩ bone marrow (58% Ϯ 12%; n ϭ 8; P Ͻ .001)
were obtained from the Jackson Laboratory. All mice were housed under
(Figure 2A). In addition, a significant decrease in the frequency of
microisolator conditions in autoclaved cages and were maintained on
CD4 single-positive cells was observed in recipients of AtmϪ/Ϫ
irradiated feed and autoclaved acidified drinking water. All sentinel mice
bone marrow (12% Ϯ 3%; n ϭ 8; P Ͻ .001) when compared with
housed in the same colony were free of viral antibodies. Four- to
recipients of Atmϩ/ϩ bone marrow (26% Ϯ 6%; n ϭ 8). The
6-week-old mice were used in all experiments.
absolute number of CD4 T cells was also significantly decreased(0.6 Ϯ 0.4 ϫ 107) when compared with recipients of Atmϩ/ϩ bone
Bone marrow transplantation
marrow (2.2 Ϯ 0.8 ϫ 107; P Ͻ .001) (Figure 2B). Similarly, thefrequency (4% Ϯ 1% vs 13% Ϯ 5%; n ϭ 8; P Ͻ .001) (Figure 2A)
Conditioning by lethal irradiation was performed as described.18 Mice
and absolute number (0.2 Ϯ 0.1 ϫ 107 vs 1.1 Ϯ 0.6 ϫ 107;
undergoing nonmyeloablative conditioning received 0.5 mg anti-CD4
P Ͻ .001) of CD8 single-positive T cells was significantly de-
antibody (GK1.5)19 and 1 mg anti-CD8 antibody (2.43)20 7 days before
creased in recipients of AtmϪ/Ϫ bone marrow when compared with
BMT and then a second dose of each antibody, together with 200 mg/kgcyclophosphamide (Cytoxan; Bristol-Myers Squibb, Princeton, NJ), 1 day
recipients of Atmϩ/ϩ bone marrow (Figure 2B). An increase in the
before BMT. Bone marrow cells were harvested from untreated donors on
frequency of CD4ϩCD8ϩ double-positive thymocytes was also
the day of BMT and were injected intravenously into conditioned recipients.
observed in recipients of AtmϪ/Ϫ bone marrow (70% Ϯ 2%; n ϭ 4)22 weeks after BMT when compared with the frequency observedin recipients of Atmϩ/ϩ bone marrow (66% Ϯ 2%; n ϭ 4; P ϭ .03)
Skin grafts
Tail skin grafting was performed as previously described.21
Flow cytometry
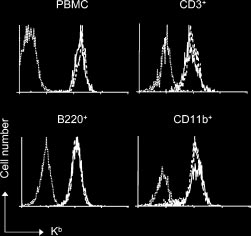
Flow cytometry was performed after gating on live cells as previouslydescribed.22 Cy-chrome conjugated anti-CD4 (RM4-5), phycoerythrin(PE)–conjugated anti-CD8 (53-6.7), fluorescein isothiocyanate (FITC)–conjugated anti–H-2Kb (AF6-88.5), anti–H-2Kk (36-7-5), anti-Ly6C (AL-21), anti-CD44, PE-conjugated anti-B220 (RA3-6B2), anti-CD122 (TM-B1), anti-CD3, and anti-CD11b were obtained from PharMingen (SanDiego, CA). Figure 1. Engraftment of either Atm؊/؊ or Atm؉/؉ donor bone marrow in conditioned recipients results in stable multilineage chimerism. Lethally irradi- Statistics
ated C3H mice were reconstituted with 107 bone marrow cells from either AtmϪ/Ϫ(solid line; n ϭ 6) mutant mice or wild-type littermate controls (dashed line; n ϭ 6).
All statistical calculations were performed using GraphPad Prism 2.01
Six weeks after BMT, PBMCs were stained with donor-specific anti–H-2Kb antibodies
software (GraphPad Software, San Diego CA). The Kaplan and Meier
and analyzed by flow cytometry. Twenty-two weeks after transplantation, blood cellswere stained with donor-specific anti–H-2Kb and lineage-specific antibodies and
method with a 95% confidence interval was used for the calculation of
were analyzed by flow cytometry for the presence of donor-derived CD3ϩ, B220ϩ, or
survival curves. Comparison of survival curves was performed using the
CD11bϩ after gating. In all experiments, PBMCs from untreated C3H mice were used
log rank test. Two-tailed t tests were used for all other statistics.
BLOOD, 15 JULY 2004 ⅐ VOLUME 104, NUMBER 2
cyclophosphamide before reconstitution with 108 C3H bone mar-row cells. Ten weeks after BMT, 7 of 9 AtmϪ/Ϫ recipients of C3Hbone marrow exhibited full donor-type multi-hematopoietic cell-lineage chimerism (Figure 3A). In contrast, none of the wild-typelittermates receiving the same preparative regimen became en-grafted with C3H-derived bone marrow cells (Figure 3A). TreatingAtmϪ/Ϫ mice with a depleting dose of anti-CD4 and anti-CD8antibodies alone was insufficient to establish engraftment of C3Hbone marrow (data not shown). Analysis of donor-derived periph-eral blood mononuclear cells (PBMCs) 52 weeks after transplanta-tion indicated that chimerism in AtmϪ/Ϫ recipients was stable(Figure 3B), demonstrating that the AtmϪ/Ϫ hematopoietic compart-ment was completely replaced with C3H-derived cells. No symp-toms of graft-versus-host disease were observed. Similar resultswere obtained using lower bone marrow doses (107-5 ϫ 108; datanot shown). ATM-deficient thymic microenvironment is able to support Figure 2. Defects in lymphocyte development observed in Atm؊/؊ mice are normal development of wild-type T cells stem cell intrinsic. (A) At 8 and 22 weeks after BMT, the thymi of C3H mice that had received either ATMϪ/Ϫ (ATMϪ/Ϫ 3 C3H) or wild-type littermate control bone marrow
Analysis of T-cell development in AtmϪ/Ϫ mice that were engrafted
cells (ATMϩ/ϩ 3 C3H) were stained with anti-CD4 and anti-CD8 antibodies and were
with C3H bone marrow revealed that the frequency of CD4ϩCD8ϩ
analyzed by flow cytometry. Shown is the frequency of each thymocyte subset inrepresentative mice. (B) Eight and 22 weeks after BMT, the total number of cells in the
double-positive thymocytes (72% Ϯ 10%; P ϭ .006; n ϭ 7) was
thymi of C3H mice that had received either AtmϪ/Ϫ (Ⅺ) or wild-type littermate control
reduced compared with that observed in AtmϪ/Ϫ mice receiving
(f) bone marrow cells were counted, and the absolute number of each population
was calculated based on the frequency of subsets as determined by flow cytometry.
Ϯ 6%; P ϭ .006; n ϭ 6) (Figure 4A). The
Shown are the combined mean and standard deviation of 3 experiments. Lethally
frequency of CD4ϩCD8ϩ double-positive thymocytes was indistin-
irradiated wild-type mice reconstituted with Atmϩ/ϩ bone marrow showed a de-
guishable from the frequency of CD4ϩCD8ϩ double-positive
creased number of CD4ϩCD8ϩ double-positive and an increased number ofsingle-positive thymocytes in comparison with untreated controls, which, based on
thymocytes observed in C3H mice (77% Ϯ 2%; n ϭ 8; P ϭ .17).
our experience, is most likely a result of damage caused by the radiation used to
Furthermore, we observed a significantly higher frequency of
condition these animals, as has been observed in previous studies.23-25
CD4ϩ (18% Ϯ 7%; n ϭ 7; P ϭ .003) and CD8ϩ (6% Ϯ 2%;
(Figure 2A). In addition, at 22 weeks, a significant decrease in thefrequency of CD4 single-positive cells was observed in recipientsof AtmϪ/Ϫ bone marrow (14% Ϯ 1%; n ϭ 4; P Ͻ .001) whencompared with recipients of Atmϩ/ϩ bone marrow (20% Ϯ 1%;n ϭ 4). The absolute number of CD4 T cells was also significantlydecreased (0.17 Ϯ 0.06 ϫ 107) when compared with recipients ofAtmϩ/ϩ bone marrow (0.5 Ϯ 0.1 ϫ 107; P Ͻ .01) (Figure 2B). Similarly, the frequency (4% Ϯ 1% vs 7% Ϯ 1%; n ϭ 4; P Ͻ .01)(Figure 2A) and absolute number (0.04 Ϯ 0.2 ϫ 107 vs0.18 Ϯ 0.06 ϫ 107; P Ͻ .01) of CD8 single-positive T cells wassignificantly decreased in recipients of AtmϪ/Ϫ bone marrow whencompared with recipients of Atmϩ/ϩ bone marrow (Figure 2B). These data suggest that the phenotypic differences in thymopoiesisobserved in recipients of AtmϪ/Ϫ bone marrow were not caused bydifferences in engraftment kinetics and that wild-type recipients ofAtm-deficient bone marrow cells display defects in T-cell develop-ment that are similar to those observed in AtmϪ/Ϫ mice. ATM deficiency decreases host resistance to bone marrow engraftment and obviates the need for irradiation Figure 3. Atm؊/؊ mice are more sensitive to conditioning than wild-type
To further analyze the ability of AtmϪ/Ϫ mice to support the
littermate controls. AtmϪ/Ϫ and Atmϩ/ϩ mice were conditioned with cyclophospha-
engraftment and development of wild-type HSCs and their prog-
mide, anti-CD4, and anti-CD8 monoclonal antibodies and were injected with 108 C3H
eny, we analyzed the engraftment of Atmϩ/ϩ bone marrow in Atm
bone marrow cells. Sixteen weeks after BMT, PBMCs were analyzed for the presenceof C3H-derived H-2Kk– or host-derived H-2b–positive cells by flow cytometry. (A)
knockout mice. Because AtmϪ/Ϫ mice are extremely sensitive to
Shown are representative examples of mice 16 weeks after BMT from 1 of 3
irradiation,9 we first set out to develop a host preparative regimen
independent experiments. Wild-type Atmϩ/ϩ mice did not show the presence of
that would not require irradiation to achieve engraftment of
donor-derived cells in PBMCs 16 weeks after transplantation. Although most (7 of 9)
wild-type donor bone marrow. AtmϪ/Ϫ and wild-type littermate
AtmϪ/Ϫ animals became fully chimeric with more than 99% donor-derived PBMCs(Chimeric), a few (2 of 9) showed no donor-derived cells (Nonchimeric). (B) Shown
mice were treated with a depleting dose of anti-CD4 and anti-CD8
are representative examples of mice 52 weeks after BMT. Note that none of the
antibodies (described in “Materials and methods”) and 200 mg/kg
AtmϪ/Ϫ mice that failed to become chimeric survived to 52 weeks.
REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
higher than the frequency of CD8ϩ thymocytes in C3H controlmice (3% Ϯ 1%; n ϭ 8; P Ͻ .001). When total cell numbers wereanalyzed, AtmϪ/Ϫ mice engrafted with C3H bone marrow hadsignificantly more CD4 (8.0 Ϯ 6.5 ϫ 106; P ϭ .03) and CD8(2.6 Ϯ 1.4 ϫ 106; P ϭ .03) single-positive thymocytes than didconditioned AtmϪ/Ϫ controls (1.7 Ϯ 0.8 ϫ 106 and 1.0 Ϯ 0.6 ϫ 106,respectively) (Figure 4B). These data suggest that replacing thebone marrow compartment of Atm-deficient mice throughtransplantation overcomes abnormalities in thymocyte subsetfrequencies observed in AtmϪ/Ϫ mice. In addition, these datasupport the hypothesis that deficiencies in T-cell developmentcaused by mutations in Atm are the result of HSC intrinsicdefects rather than defects in the microenvironment in which theprogeny of these cells mature. Improved T-cell development in the thymi of Atm؊/؊ mice reconstituted with C3H bone marrow transplants results in increased frequency of T cells in peripheral blood
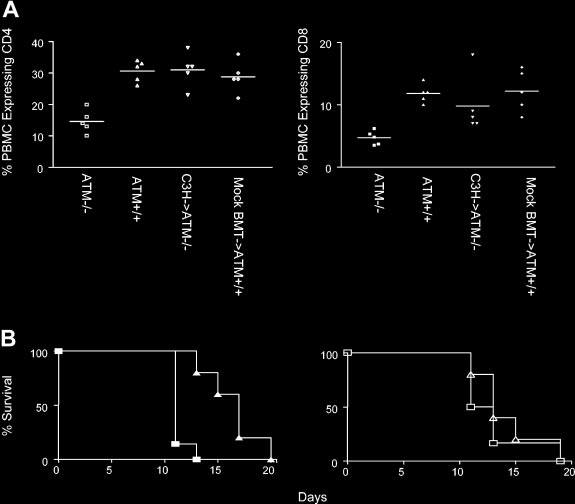
The frequency of CD4 T cells in the blood of AtmϪ/Ϫ mutant mice
Figure 4. T-cell development is normal in Atm؊/؊ mutant mice that receive C3H
(15% Ϯ 4%; n ϭ 5) is significantly lower than in wild-type mice
bone marrow cells. (A) AtmϪ/Ϫ mice were treated with anti-CD4 and anti-CD8
(30% Ϯ 3%; n ϭ 5; P Ͻ .001 Figure 5), most likely because of
antibodies and cyclophosphamide before receiving 108 C3H bone marrow cells. Twelve weeks after BMT, mice were killed and thymi were analyzed by flow cytometry
poor thymic output in AtmϪ/Ϫ mice, as suggested previously.9 In
after cell surface staining. Shown is a flow cytometry profile from representative mice.
contrast, the frequency of CD4 T cells in the blood of AtmϪ/Ϫ
(B) The total number of cells in each thymus was counted, and the absolute number
mutant mice reconstituted with C3H bone marrow (31% Ϯ 5%;
of each thymocyte subset was calculated based on the frequency of each subset as
determined by flow cytometry. Shown is the absolute number of cells in each
ϭ 5) was the same as the frequency of CD4 T cells in the blood of
thymocyte subset in AtmϪ/Ϫ mice that were engrafted with C3H bone marrow (f) and
Atmϩ/ϩ controls (29% Ϯ 5%; n ϭ 5; P ϭ .5) that were treated with
conditioned control AtmϪ/Ϫ mice (Ⅺ). Shown are the combined mean and standard
anti–T-cell antibodies and cyclophosphamide and were injected
with 108 C3H bone marrow cells, as described (mock BMTcontrols), conditioning that does not allow the engraftment of
n ϭ 7; P ϭ .02) single-positive thymocytes in AtmϪ/Ϫ mice recon-
donor-derived cells. When compared with unmanipulated controls,
stituted with C3H bone marrow when compared with AtmϪ/Ϫ mice
the frequency of CD4 T cells in the blood of AtmϪ/Ϫ mutant mice
that received conditioning alone (6% Ϯ 3% and 4% Ϯ 2%, respec-
that received C3H bone marrow was significantly higher than the
tively; n ϭ 6). The frequency of single-positive CD4 T cells in the
frequency of CD4 T cells in the blood of untreated AtmϪ/Ϫ mutant
thymi of AtmϪ/Ϫ mice engrafted with C3H bone marrow was the
mice (P ϭ .0005), but it was not significantly different than the
same as that observed in untreated C3H controls (13% Ϯ 4%;
frequency of CD4 T cells found in the blood of untreated wild-type
n ϭ 8; P ϭ .15). The frequency of CD8ϩ thymocytes (6% Ϯ 2%;
littermates (P ϭ .9). Similar results were observed for the fre-
n ϭ 7) in AtmϪ/Ϫ mice engrafted with C3H bone marrow was
quency of CD8 T cells in the blood (Figure 5A). The frequency of
Figure 5. Restoration of lymphocyte numbers and immune function in Atm؊/؊ mutant mice reconstituted with C3H bone marrow cells. (A) Left panel: the frequency of CD4 T cells (left panel) in PBMCs of AtmϪ/Ϫ mice (Ⅺ), Atmϩ/ϩ mice (‚), AtmϪ/Ϫ mice reconstituted with C3H bone marrow (ƒ), and wild-type littermate controls receiving the BMT regimen (mock BMT, छ). Right panel: the frequency of CD8 T cells in PBMCs of AtmϪ/Ϫ mice (f), Atmϩ/ϩ mice (Œ), AtmϪ/Ϫ mice receiving C3H bone marrow (), and wild-type littermate controls receiving the BMT regimen (mock BMT, ࡗ). Horizontal bars indicate arithmetic mean. (B) Left panel: rejection of B6.CH-2bm1 skin grafts by unmodified AtmϪ/Ϫ recipients (Œ) and Atmϩ/ϩ littermates (f). Right panel: rejection of B6.CH-2bm1 skin graft by AtmϪ/Ϫ mice that received C3H transplanted bone marrow (‚) and mock BMT Atmϩ/ϩ controls (Ⅺ). Shown are the results of 1 of 2 experiments.
BLOOD, 15 JULY 2004 ⅐ VOLUME 104, NUMBER 2
CD8 T cells in the blood of AtmϪ/Ϫ mutant mice (5% Ϯ 1%; n ϭ 5)
early of thymic lymphoma.9 AtmϪ/Ϫ mice reconstituted with
is significantly lower than in wild-type mice (12% Ϯ 1%; n ϭ 5;
wild-type C3H bone marrow had a significantly lower frequency of
P Ͻ .001) (Figure 5). In contrast, the frequency of CD8 T cells in
CD44hi CD8 T cells (39% Ϯ 10%; n ϭ 5) than unmanipulated
the blood of AtmϪ/Ϫ mutant mice reconstituted with C3H bone
AtmϪ/Ϫ mice (65% Ϯ 8%; n ϭ 5; P ϭ .0004). The frequency of
marrow (10% Ϯ 5%; n ϭ 5) was the same as the frequency of CD8
CD44hi CD8 T cells in AtmϪ/Ϫ mice reconstituted with C3H bone
T cells in the blood of mock BMT Atmϩ/ϩ controls (12% Ϯ 3%;
marrow did not differ from that of mock BMT Atmϩ/ϩ control mice
n ϭ 5; P ϭ .4). When compared with unmanipulated controls, the
(34% Ϯ 6%; n ϭ 5; P ϭ .4). Similarly, AtmϪ/Ϫ mice reconstituted
frequency of CD8 T cells in the blood of AtmϪ/Ϫ mutant mice that
with C3H bone marrow had a significantly lower frequency of
received C3H bone marrow was significantly higher than the
CD122/Ly6C double-positive CD8 T cells (10% Ϯ 5%; n ϭ 5;
frequency of CD4 T cells in the blood of untreated AtmϪ/Ϫ mutant
P Ͻ .001) than AtmϪ/Ϫ controls (41% Ϯ 11%; n ϭ 5), and the
mice (P ϭ .04) but was not significantly different than the fre-
frequency of these cells did not differ significantly from that of
quency of CD4 T cells found in the blood of untreated wild-type
wild-type littermates that received mock BMT (5% Ϯ 2%; n ϭ 5;
littermates (P ϭ .4). Thus, replacing the AtmϪ/Ϫ hematopoietic
P ϭ .06). The frequency of CD44hi CD4 T cells in the blood of
compartment by transplanting wild-type bone marrow overcomes
AtmϪ/Ϫ mice that received C3H bone marrow transplants was also
deficiencies in thymocyte development and overcomes the de-
significantly lower (11% Ϯ 1%; n ϭ 5) than the frequency ob-
crease in peripheral T-cell numbers observed in AtmϪ/Ϫ mice.
served in AtmϪ/Ϫ controls (24% Ϯ 5%; n ϭ 5; P Ͻ .001). Thefrequency of CD44hi CD4 T cells in AtmϪ/Ϫ mice reconstituted withC3H bone marrow did not differ significantly from that of
Transplanting wild-type bone marrow into Atm؊/؊ mice results
wild-type littermate mice that received mock BMT (16% Ϯ 6%;
in normal memory T-cell frequencies
n ϭ 5; P ϭ .055). These data suggest that the frequency of CD8
Patients with A-T have increased frequencies of memory T cells in
and CD4 memory T cells in AtmϪ/Ϫ mutant mice was restored to
their blood and reduced numbers of naive T cells.14 To determine
normal after replacement of the hematopoietic compartment by
whether this was also true of AtmϪ/Ϫ mutant mice, PBMCs from 4-
transplantation of wild-type bone marrow.
to 6-week-old AtmϪ/Ϫ and wild-type littermates were analyzed bycell surface staining and flow cytometry for expression markers on
Restoring immune function in Atm؊/؊ mice after BMT
memory T cells. Memory CD8 T cells are characterized by cellsurface expression of CD122, CD44, and Ly6C.26 As observed in
To determine whether replacing the hematopoietic compartment in
A-T patients, the frequency of CD122ϩ, Ly6Cϩ CD8 T cells was
AtmϪ/Ϫ mice can overcome immunoincompetence, we compared
significantly higher in AtmϪ/Ϫ mutant mice (41% Ϯ 11%; n ϭ 5)
the ability of AtmϪ/Ϫ mice reconstituted with wild-type C3H bone
than in wild-type littermate controls (5% Ϯ 1%; n ϭ 5; P Ͻ .001)
marrow and Atmϩ/ϩ controls to reject skin allografts. Unmanipu-
(Table 1). Similarly, the frequency of CD44hi CD8ϩ T cells was
lated AtmϪ/Ϫ mice (H-2b) exhibited delayed rejection of skin
significantly higher in AtmϪ/Ϫ mice (65% Ϯ 8%; n ϭ 5) than
allografts from allogeneic B6.CH-2bm1 mice (median survival time
wild-type controls (24% Ϯ 2%; n ϭ 5; P Ͻ .001) (Table 1). The
[MST], 17 days; n ϭ 7) when compared with healthy littermate
frequency of CD44hi CD4ϩ T cells in AtmϪ/Ϫ mice was also
Atmϩ/ϩ controls (MST, 11 days; n ϭ 5; P ϭ .002) (Figure 5B).
significantly higher (24% Ϯ 5%; n ϭ 5) than in Atmϩ/ϩ mice
Therefore, as observed in humans, Atm deficiency leads to
(7% Ϯ 2%; n ϭ 5; P Ͻ .001). Thus, the frequency of T cells
hyporesponsiveness to alloantigen.3 In contrast, AtmϪ/Ϫ mice
expressing memory markers is increased in AtmϪ/Ϫ mice compared
reconstituted with wild-type C3H bone marrow were able to reject
with healthy controls, as is observed in A-T patients.
B6.CH-2bm1 skin allografts with the same kinetics (MST, 13; n ϭ 5;
To determine whether the altered memory-like T-cell phenotype
P ϭ .52) observed for Atmϩ/ϩ mice receiving mock BMT (MST, 12
is overcome in mice that receive bone marrow transplants from
days; n ϭ 6) (Figure 5B). The median survival time of B6.CH-2bm1
wild-type mice, AtmϪ/Ϫ mutant mice or wild-type littermate mock
on AtmϪ/Ϫ mice reconstituted with wild-type C3H bone marrow
BMT controls were reconstituted, as described, with C3H bone
was the same as that observed for unmanipulated healthy littermate
marrow. Twenty-five weeks after transplantation, the frequency of
Atmϩ/ϩ controls (P Ͼ .05). These data suggest that replacing the
memory marker–positive T cells in the blood of AtmϪ/Ϫ mice
AtmϪ/Ϫ hematopoietic compartment through BMT can overcome
receiving transplanted bone marrow was compared with the
the immunodeficiency observed in AtmϪ/Ϫ mice.
frequency observed in 4- to 6-week-old AtmϪ/Ϫ mice and in Atmϩ/ϩand mock BMT Atmϩ/ϩ control mice. It was not possible to use
Replacing the bone marrow compartment in Atm؊/؊ mice
age-matched AtmϪ/Ϫ mutant mice because these mice die relatively
prevents the generation of thymic lymphoma AtmϪ/Ϫ mutant mice acquire fatal thymic malignancies as early as 9
Table 1. Expression of memory phenotype markers in PBMCs
weeks of age, and, by 20 weeks of age, essentially all AtmϪ/Ϫ
Memory CD4
mutant mice develop thymic lymphomas that prove fatal by 30
Memory CD8 T cells
weeks of age.9 To determine whether replacing the bone marrow
CD44؉CD4؉, CD44؉CD8؉, CD122؉Ly6C؉CD8؉,
compartment in AtmϪ/Ϫ mice through BMT could delay or prevent
the development of thymic lymphomas, 4- to 6-week-old AtmϪ/Ϫ
mutant mice were conditioned and reconstituted as described with
108 C3H bone marrow cells. Animals were monitored long term for
survival. As expected, AtmϪ/Ϫ mice that underwent conditioning
alone acquired thymic lymphoma and were killed (MST, 11.5
Values presented in the table are means and standard deviations.
weeks; range, 6-22 weeks after transplantation; n ϭ 12). In con-
*Percentage of CD4 and CD8 T cells that express CD44.
trast, AtmϪ/Ϫ mice reconstituted with C3H bone marrow displayed
†Percentage of CD8 T cells that express CD122 and Ly6C. Results from 1 of 2
experiments are shown; n ϭ 5 for all groups.
prolonged survival (MST, more than 60 weeks; n ϭ 21; P Ͻ .001)
REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
overcome, resulting in normal responses to alloantigen based onthe rejection of skin allografts. These data suggest that BMT canbe used to overcome defects in T-cell development that lead toimmunodeficiency in Atm-deficient mice.
In AtmϪ/Ϫ mice, nonmyeloablative conditioning consisting of
T-cell depletion and cyclophosphamide administration wassufficient to induce full donor-type chimerism. However, thesame preparative regimen failed to induce donor-type chimer-ism in wild-type mice. These data suggest that barriers to bone
Figure 6. Replacement of the Atm؊/؊ hematopoietic compartment by BMT
marrow engraftment are significantly reduced in AtmϪ/Ϫ mice.27
prevents lymphoma. AtmϪ/Ϫ mice were conditioned with cyclophosphamide and
It is possible that reduced barriers to the engraftment of
anti-CD4 and -CD8 monoclonal antibodies. Mice that received C3H bone marrow (Œ)
donor-type bone marrow may reflect a competitive disadvantage
had a significantly longer lifespans than AtmϪ/Ϫ control mice that did not receive C3Hbone marrow (f). Shown are the combined results of 5 experiments.
of AtmϪ/Ϫ bone marrow as a result of cell-intrinsic defects. As aresult, these cells may be unable to compete with wild-type cellsfor bone marrow niches. Alternatively, Atm deficiency may
(Figure 6). In this group, only 1 mouse was confirmed to have died
increase sensitivity to the immunosuppressive effects of cyclo-
of thymic lymphoma 13 weeks after transplantation based on
phosphamide, which, in turn, may allow donor bone marrow to
postmortem examination. We were unable to detect thymic lym-
engraft more efficiently by reducing antidonor immune re-
phoma in the 2 mice in this group that died at 30 and 43 weeks after
sponses more effectively than in wild-type mice. Patients with
BMT. The remaining mice survived more than 52 weeks after BMT
A-T appear to be more susceptible to adverse effects from
or were killed at earlier time points without evidence of thymic
agents such as cyclophosphamide.5 Previous work has suggested
lymphoma. We did not inject bone marrow cells from untreated
that host T-cell depletion is critical for efficient bone marrow
AtmϪ/Ϫ animals into control mice because it was possible that
engraftment.22,28,29 Insofar as AtmϪ/Ϫ mice exhibit reduced
transferring malignant cells from the untreated AtmϪ/Ϫ donorscould artificially accelerate deaths in the control population. These
numbers of mature T cells, it is also possible that immunodefi-
data suggest that replacing the AtmϪ/Ϫ hematopoietic compartment
ciency observed in AtmϪ/Ϫ mice may reduce the requirement for
through BMT prevents the development of thymic lymphoma.
rigorous myeloablation to achieve donor bone marrow engraft-ment. Regardless of the mechanism that allows for full replace-ment of the hematopoietic compartment in AtmϪ/Ϫ mice usingrelatively mild host conditioning, our results suggest that similar
Discussion
defects in humans may make it possible to achieve fulldonor-type chimerism with minimal conditioning. Although it
Immunodeficiencies can arise from defects in hematopoietic
remains to be determined how patients with A-T will be able to
stem cells that give rise to the cells of the immune system or
tolerate cytoreductive drugs, cyclophosphamide is routinely
from defects in the microenvironment in which immune system
used in patients undergoing BMT.30-34 In addition, several
cells mature, such as the thymus. Our data demonstrate that
clinical protocols using human-specific T-cell depletion and
AtmϪ/Ϫ bone marrow contains T-cell progenitors that give rise
cyclophosphamide have been shown to be tolerated in humans35;
to thymic precursors unable to develop normally into mature
therefore, we suggest that it may be possible to develop similar
single-positive T cells in a normal thymic environment. Wild-
conditioning regimens that will be clinically relevant. The
type mice reconstituted with AtmϪ/Ϫ bone marrow exhibit
ability to achieve full donor chimerism using a relatively
a block in thymocyte development similar to that observed
nontoxic host-conditioning regimen would make BMT a clini-
in AtmϪ/Ϫ mice, indicating that the defects in T-cell develop-
cally acceptable means to address hematologic defects associ-
ment observed in these mice are not solely the result of an
abnormal thymic microenvironment. Despite the reported abnor-
Thymic lymphomas have been shown to occur at a high
malities in T-cell development observed in the thymi of AtmϪ/Ϫ
frequency in Atm-deficient mice, resulting in death by 30 weeks
mice, progeny of wild-type bone marrow cells were able to
of age. Replacing the hematopoietic system in AtmϪ/Ϫ mutant
develop normally and to restore normal T-cell development in
mice through BMT prevented the occurrence of thymic lympho-
the thymi of AtmϪ/Ϫ mice. Together, these data suggest that
mas and resulted in a significantly prolonged lifespan that was
immunodeficiencies observed in AtmϪ/Ϫ mice are attributable to
identical to that observed for healthy controls. Although the role
intrinsic defects in the progeny of bone marrow–derived cellsrather than to the microenvironment in which these cells
of antigen receptor gene rearrangement in the generation of
lymphoma in AtmϪ/Ϫ is controversial,36-38 our results strongly
Patients with A-T have an increased frequency of memory T
suggest that in AtmϪ/Ϫ mice, essentially all malignancies
cells in the blood.14 We were able to demonstrate a similar defect
observed are hematologic in origin and that replacing the
in AtmϪ/Ϫ mice. BMT was able to restore the frequency of
Atm-deficient bone marrow compartment prevents their occur-
memory T cells in the periphery of AtmϪ/Ϫ mice to levels
rence. Although the occurrence of malignancy in AtmϪ/Ϫ mice
observed in healthy controls. Furthermore, replacing the hema-
was prevented by inducing full donor-type chimerism, it is
topoietic compartment in AtmϪ/Ϫ mice by transplanting wild-
unclear whether full donor-type chimerism is necessary to
type bone marrow restored the frequency of mature CD4 and
reduce the occurrence of lymphoma. We are investigating the
CD8 T cells in the peripheral blood to normal. Replacing the
level of donor-type chimerism needed to achieve significant
AtmϪ/Ϫ hematopoietic compartment through BMT also allowed
protection from lymphoma and are determining whether solid
the functional immunodeficiency observed in AtmϪ/Ϫ mice to be
tumors develop in these mice as they age.
BLOOD, 15 JULY 2004 ⅐ VOLUME 104, NUMBER 2
A significant proportion of A-T patients experience recurrent
pulmonary infections resulting from immunodeficiency. Hemato-
Acknowledgments
logic malignancies occur in as many as 40% of patients4 and,together with bronchial infection, are the major causes of death inA-T patients. Demonstrating that BMT may overcome immune
We thank Lee Bar-Sagi for expert technical assistance and Jessica
system defects and the occurrence of hematologic malignancy in
Sheehan for secretarial support. We thank Drs David H. Sachs,
AtmϪ/Ϫ mice opens up the possibility that similar therapies may
Megan Sykes, and Ronjon Chakraverty for critical review of the
eventually be able to alleviate these major causes of morbidity and
manuscript. We also thank Brad Margus for his support and
References
1. Swift M. Genetics and epidemiology of ataxia-
15. Buckley RH. Primary cellular immunodeficien-
mixed chimeras established with a cyclophospha-
telangiectasia. Kroc Found Ser. 1985;19:133-
cies. J Allergy Clin Immunol. 2002;109:747-757.
mide-based nonmyeloablative conditioning regi-
16. Saha K, Chopra K. Primary immune disorders in
men. Biol Blood Marrow Transplant. 1999;5:133-
2. Swift M, Morrell D, Cromartie E, et al. The inci-
children and their diagnosis. J Commun Dis.
dence and gene frequency of ataxia-telangiecta-
28. Sykes M, Sheard MA, Sachs DH. Effects of T cell
sia in the United States. Am J Hum Genet. 1986;
17. Micheli R, Pirovano S, Calandra G, et al. Low thy-
depletion in radiation bone marrow chimeras, II:
mic output and reduced heterogeneity of alpha/
requirement for allogeneic T cells in the reconsti-
3. Lavin MF, Shiloh Y. The genetic defect in ataxia-
beta, but not gamma/delta, T lymphocytes in in-
tuting bone marrow inoculum for subsequent re-
telangiectasia. Annu Rev Immunol. 1997;15:177-
fants with ataxia-telangiectasia. Neuropediatrics.
sistance to breaking of tolerance. J Exp Med.
4. Morrell D, Cromartie E, Swift M. Mortality and
18. Bagley J, Tian C, Sachs DH, Iacomini J. Induction
29. Sykes M, Sheard M, Sachs DH. Effects of T cell
cancer incidence in 263 patients with ataxia-tel-
of T-cell tolerance to an MHC class I alloantigen
depletion in radiation bone marrow chimeras, I:
angiectasia. J Natl Cancer Inst. 1986;77:89-92.
by gene therapy. Blood. 2002;99:4394-4399.
evidence for a donor cell population which in-
5. Sandoval C, Swift M. Treatment of lymphoid ma-
19. Dialynas DP, Quan ZS, Wall KA, et al. Character-
creases allogeneic chimerism but which lacks the
lignancies in patients with ataxia-telangiectasia.
ization of the murine T cell surface molecule des-
potential to produce GVHD. J Immunol. 1988;
ignated L3T4, identified by monoclonal antibody
6. Abadir R, Hakami N. Ataxia telangiectasia with
GK1.5: similarity of L3T4 to the human Leu 3/T4
30. Thomas ED, Sanders JE, Flournoy N, et al. Mar-
cancer: an indication for reduced radiotherapy
molecule. J Immunol. 1984;131:2445-2451.
row transplantation for patients with acute lym-
and chemotherapy doses. Br J Radiol. 1983;56:
20. Sarmiento M, Glasebrook AL, Fitch FW. IgG or
phoblastic leukemia: a long-term follow-up.
IgM monoclonal antibodies reactive with different
7. Weyl BA, Rosenthal J, Dale J, et al. Ataxia telan-
determinants on the molecular complex bearing
31. Thomas ED, Buckner CD, Clift RA, et al. Marrow
giectasia and lymphoma: an indication for indi-
Lyt2 antigen block T cell-mediated cytolysis in the
transplantation for acute nonlymphoblastic leuke-
vidualized chemotherapy dosing—report of treat-
absence of complement. J Immunol. 1980;125:
mia in first remission. N Engl J Med. 1979;301:
ment in a highly inbred Arab family. Pediatr
21. Ildstad ST, Wren SM, Bluestone JA, Barbieri SA,
8. Borghesani PR, Alt FW, Bottaro A, et al. Abnormal
Sachs DH. Characterization of mixed allogeneic
32. Santos GW. Marrow transplantation in acute non-
development of Purkinje cells and lymphocytes in
chimeras: immunocompetence, in vitro reactivity,
lymphocytic leukemia. Blood. 1989;74:901-908.
Atm mutant mice. Proc Natl Acad Sci U S A.
and genetic specificity of tolerance. J Exp Med.
33. Beutler E, Blume KG, Bross KJ, et al. Bone mar-
row transplantation as the treatment of choice for
9. Barlow C, Hirotsune S, Paylor R, et al. Atm-defi-
22. Bagley J, Tian C, Sachs DH, Iacomini J. T cells
“good risk” adult patients with acute leukemia.
cient mice: a paradigm of ataxia telangiectasia.
mediate resistance to genetically modified bone
Trans Assoc Am Physicians. 1979;92:189-195.
marrow in lethally irradiated recipients. Trans-
34. Spitzer TR, McAfee SL, Dey BR, et al. Nonmy-
10. Xu Y, Ashley T, Brainerd EE, et al. Targeted dis-
eloablative haploidentical stem-cell transplanta-
ruption of ATM leads to growth retardation, chro-
23. Chung B, Barbara-Burnham L, Barsky L, Wein-
tion using anti-CD2 monoclonal antibody (MEDI-
mosomal fragmentation during meiosis, immune
berg K. Radiosensitivity of thymic interleukin-7
507)-based conditioning for refractory hematologic
defects and thymic lymphoma. Genes Dev. 1996;
production and thymopoiesis after bone marrow
malignancies. Transplantation. 2003;75:1748-1751.
transplantation. Blood. 2001;98:1601-1606.
35. Kim HJ, Park CY, Park YH, et al. Successful allo-
11. Elson A, Wang Y, Daugherty CJ, et al. Pleiotropic
24. Bolotin E, Smogorzewska M, Smith S, Widmer M,
geneic hematopoietic stem cell transplantation
defects in ataxia-telangiectasia protein-deficient
Weinberg K. Enhancement of thymopoiesis after
using triple agent immunosuppression in severe
mice. Proc Natl Acad Sci U S A. 1996;93:13084-
bone marrow transplant by in vivo interleukin-7.
aplastic anemia patients. Bone Marrow Trans-
12. Giovannetti A, Mazzetta F, Caprini E, et al.
25. Min D, Taylor PA, Panoskaltsis-Mortari A, et al.
36. Liao MJ, Van Dyke T. Critical role for Atm in sup-
Skewed T-cell receptor repertoire, decreased thy-
Protection from thymic epithelial cell injury by ker-
pressing V(D)J recombination-driven thymic lym-
mic output, and predominance of terminally differ-
atinocyte growth factor: a new approach to im-
phoma. Genes Dev. 1999;13:1246-1250.
entiated T cells in ataxia telangiectasia. Blood.
prove thymic and peripheral T-cell reconstitution
after bone marrow transplantation. Blood. 2002;
37. Petiniot LK, Weaver Z, Barlow C, et al. Recombi-
13. Schubert R, Reichenbach J, Zielen S. Deficien-
nase-activating gene (RAG) 2-mediated V(D)J
cies in CD4ϩ and CD8ϩ T cell subsets in ataxia
26. Goldrath AW, Bogatzki LY, Bevan MJ. Naive T
recombination is not essential for tumorigenesis
telangiectasia. Clin Exp Immunol. 2002;129:125-
cells transiently acquire a memory-like phenotype
in atm-deficient mice. Proc Natl Acad Sci U S A.
during homeostasis-driven proliferation. J Exp
14. Paganelli R, Scala E, Scarselli E, et al. Selective
38. Petiniot LK, Weaver Z, Vacchio M, et al. RAG-
deficiency of CD4ϩ/CD45RAϩ lymphocytes in
27. Pelot MR, Pearson DA, Swenson K, et al. Lym-
mediated V(D)J recombination is not essential for
patients with ataxia-telangiectasia. J Clin Immu-
phohematopoietic graft-vs.-host reactions can be
tumorigenesis in Atm-deficient mice. Mol Cell
induced without graft-vs.-host disease in murine
REGLAMENTO DEL SERVICIO POLICIAL DE CARRERA DEL ESTADO DE COAHUILA PUBLICADO EN EL P. O. N° DEL DE OCTUBRE DE 1999 REGLAMENTO EMITIDO DURANTE LA ADMINISTRACION DEL C. ROGELIO MONTEMAYOR SEGUY ROGELIO MONTEMAYOR SEGUY, Gobernador Constitucional del Estado de Coahuila de Zaragoza, en ejercicio de las facultades que me confieren los artículos 82, fracción XVIII y 85, párrafo tercer
Aesthestic Plastic SurgeryMaxillo-facial SurgeryFollicular hair transplantation Hair transplantation Prior to treatment: No aspirin or other medicine that thins the blood should be taken (otherwise there is anincreased tendency to haemorrhage)No sedatives are to be taken on the day of treatment. You will receive the appropriatemedication from us, if required. Please wash your hair before you
 REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
would overcome the observed hematologic abnormalities. Ourresults indicate that full donor-type hematopoiesis can be achieved
in AtmϪ/Ϫ mice using clinically relevant host conditioning, result-ing in the restoration of normal immune system function. In
Defects in lymphocyte development observed in Atm؊/؊ mice
REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
would overcome the observed hematologic abnormalities. Ourresults indicate that full donor-type hematopoiesis can be achieved
in AtmϪ/Ϫ mice using clinically relevant host conditioning, result-ing in the restoration of normal immune system function. In
Defects in lymphocyte development observed in Atm؊/؊ mice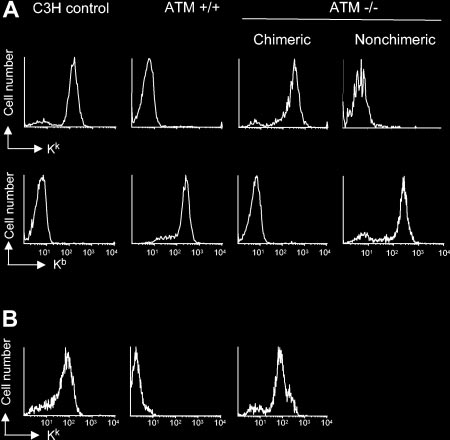
 BLOOD, 15 JULY 2004 ⅐ VOLUME 104, NUMBER 2
cyclophosphamide before reconstitution with 108 C3H bone mar-row cells. Ten weeks after BMT, 7 of 9 AtmϪ/Ϫ recipients of C3Hbone marrow exhibited full donor-type multi-hematopoietic cell-lineage chimerism (Figure 3A). In contrast, none of the wild-typelittermates receiving the same preparative regimen became en-grafted with C3H-derived bone marrow cells (Figure 3A). TreatingAtmϪ/Ϫ mice with a depleting dose of anti-CD4 and anti-CD8antibodies alone was insufficient to establish engraftment of C3Hbone marrow (data not shown). Analysis of donor-derived periph-eral blood mononuclear cells (PBMCs) 52 weeks after transplanta-tion indicated that chimerism in AtmϪ/Ϫ recipients was stable(Figure 3B), demonstrating that the AtmϪ/Ϫ hematopoietic compart-ment was completely replaced with C3H-derived cells. No symp-toms of graft-versus-host disease were observed. Similar resultswere obtained using lower bone marrow doses (107-5 ϫ 108; datanot shown).
BLOOD, 15 JULY 2004 ⅐ VOLUME 104, NUMBER 2
cyclophosphamide before reconstitution with 108 C3H bone mar-row cells. Ten weeks after BMT, 7 of 9 AtmϪ/Ϫ recipients of C3Hbone marrow exhibited full donor-type multi-hematopoietic cell-lineage chimerism (Figure 3A). In contrast, none of the wild-typelittermates receiving the same preparative regimen became en-grafted with C3H-derived bone marrow cells (Figure 3A). TreatingAtmϪ/Ϫ mice with a depleting dose of anti-CD4 and anti-CD8antibodies alone was insufficient to establish engraftment of C3Hbone marrow (data not shown). Analysis of donor-derived periph-eral blood mononuclear cells (PBMCs) 52 weeks after transplanta-tion indicated that chimerism in AtmϪ/Ϫ recipients was stable(Figure 3B), demonstrating that the AtmϪ/Ϫ hematopoietic compart-ment was completely replaced with C3H-derived cells. No symp-toms of graft-versus-host disease were observed. Similar resultswere obtained using lower bone marrow doses (107-5 ϫ 108; datanot shown).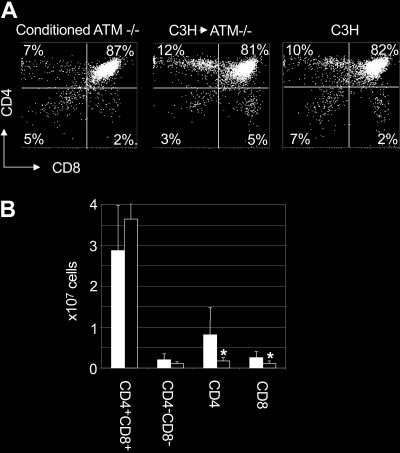
 REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
higher than the frequency of CD8ϩ thymocytes in C3H controlmice (3% Ϯ 1%; n ϭ 8; P Ͻ .001). When total cell numbers wereanalyzed, AtmϪ/Ϫ mice engrafted with C3H bone marrow hadsignificantly more CD4 (8.0 Ϯ 6.5 ϫ 106; P ϭ .03) and CD8(2.6 Ϯ 1.4 ϫ 106; P ϭ .03) single-positive thymocytes than didconditioned AtmϪ/Ϫ controls (1.7 Ϯ 0.8 ϫ 106 and 1.0 Ϯ 0.6 ϫ 106,respectively) (Figure 4B). These data suggest that replacing thebone marrow compartment of Atm-deficient mice throughtransplantation overcomes abnormalities in thymocyte subsetfrequencies observed in AtmϪ/Ϫ mice. In addition, these datasupport the hypothesis that deficiencies in T-cell developmentcaused by mutations in Atm are the result of HSC intrinsicdefects rather than defects in the microenvironment in which theprogeny of these cells mature.
REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
higher than the frequency of CD8ϩ thymocytes in C3H controlmice (3% Ϯ 1%; n ϭ 8; P Ͻ .001). When total cell numbers wereanalyzed, AtmϪ/Ϫ mice engrafted with C3H bone marrow hadsignificantly more CD4 (8.0 Ϯ 6.5 ϫ 106; P ϭ .03) and CD8(2.6 Ϯ 1.4 ϫ 106; P ϭ .03) single-positive thymocytes than didconditioned AtmϪ/Ϫ controls (1.7 Ϯ 0.8 ϫ 106 and 1.0 Ϯ 0.6 ϫ 106,respectively) (Figure 4B). These data suggest that replacing thebone marrow compartment of Atm-deficient mice throughtransplantation overcomes abnormalities in thymocyte subsetfrequencies observed in AtmϪ/Ϫ mice. In addition, these datasupport the hypothesis that deficiencies in T-cell developmentcaused by mutations in Atm are the result of HSC intrinsicdefects rather than defects in the microenvironment in which theprogeny of these cells mature.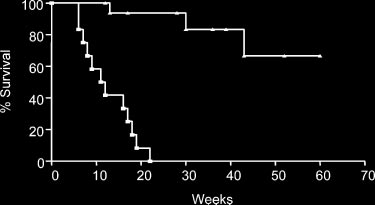 REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
overcome, resulting in normal responses to alloantigen based onthe rejection of skin allografts. These data suggest that BMT canbe used to overcome defects in T-cell development that lead toimmunodeficiency in Atm-deficient mice.
REPAIR OF T-CELL DEVELOPMENT IN AtmϪ/Ϫ MICE
overcome, resulting in normal responses to alloantigen based onthe rejection of skin allografts. These data suggest that BMT canbe used to overcome defects in T-cell development that lead toimmunodeficiency in Atm-deficient mice.